

Реферат дыхательная система у рыб: Дыхательная система у рыб 4 класс доклад
Iteach
Материал из ИнтеВики — обучающей площадкой для проведения тренингов программы Intel
|
 0/Екатеринбург сентябрь 2013 преподаватель – Ирина Нургалеева
0/Екатеринбург сентябрь 2013 преподаватель – Ирина Нургалеева
 Ноябрьск — преподаватель — Елена Ремизова
Ноябрьск — преподаватель — Елена Ремизова
 ..
.. Дыхание
Эволюция дыхательной системы
Всё живое на Земле существует за сёт солнечного тепла и энергии, достигающей поверхности нашей планеты. Все животные и человек приспособились добывать энергию из синтезированных растениями органических веществ. Чтобы использовать энергию Солнца, заключённую в молекулах органических веществ, её необходимо высвободить, окислив эти вещества. Чаще всего в качестве окислителя используют кислород воздуха, благо он составляет почти четверть объёма окружающей атмосферы.
Одноклеточные простейшие животные, кишечнополостные, свободноживущие плоские и круглые черви дышат всей поверхностью тела. Специальные органы дыхания — перистые жабры появляются у морских кольчатых червей и у водных членистоногих. Органами дыхания членистоногих являются трахеи, жабры, листовидные лёгкие расположенные в углублениях покрова тела.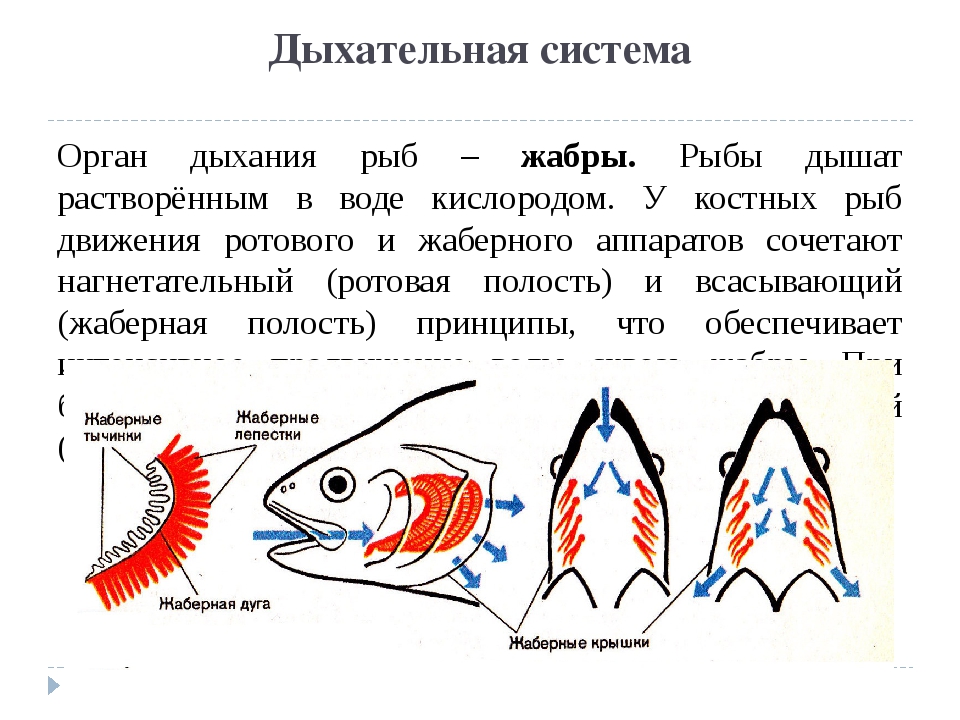 Система органов дыхания ланцетника представлена жаберными щелями, пронизывающими стенку переднего отдела кишечника — глотку. У рыб под жаберными крышками располагаются жабры, обильно пронизанными мельчайшими кровеносными сосудами. У наземных позвоночных органами дыхания являются лёгкие. Эволюция дыхания у позвоночных шла по пути увеличения площади легочных перегородок, участвующих в газообмене, совершенствования транспортных систем доставки кислорода к клеткам, расположенным внутри организма, и развития систем, обеспечивающих вентиляцию органов дыхания.
Система органов дыхания ланцетника представлена жаберными щелями, пронизывающими стенку переднего отдела кишечника — глотку. У рыб под жаберными крышками располагаются жабры, обильно пронизанными мельчайшими кровеносными сосудами. У наземных позвоночных органами дыхания являются лёгкие. Эволюция дыхания у позвоночных шла по пути увеличения площади легочных перегородок, участвующих в газообмене, совершенствования транспортных систем доставки кислорода к клеткам, расположенным внутри организма, и развития систем, обеспечивающих вентиляцию органов дыхания.
Строение и функции органов дыхания
Необходимым условием жизнедеятельности организма является постоянный газообмен между организмом и окружающей средой. Органы, по которым циркулируют вдыхаемый и выдыхаемый воздух, объединяются в дыхательный аппарат. Систему органов дыхания образуют носовая полость, глотка, гортань, трахея, бронхи и лёгкие. Большинство из них представляют собой воздухоносные пути и служат для проведения воздуха в лёгкие. В лёгких и происходят процессы газообмена. При дыхании организм получает из воздуха кислород, который разносится кровью по всему телу. Кислород участвует в сложных окислительных процессах органических веществ, при котором освобождается необходимая организму энергия. Конечные продукты распада — углекислота и частично вода — выводятся из организма в окружающую среду через органы дыхания.
В лёгких и происходят процессы газообмена. При дыхании организм получает из воздуха кислород, который разносится кровью по всему телу. Кислород участвует в сложных окислительных процессах органических веществ, при котором освобождается необходимая организму энергия. Конечные продукты распада — углекислота и частично вода — выводятся из организма в окружающую среду через органы дыхания.
| Название отдела | Особенности строения | Функции |
| Воздухоносные пути | ||
| Полость носа и носоглотка | Извилистые носовые ходы. Слизистая снабжена капиллярами, покрыта мерцательным эпителием и имеет много слизистых железок. Есть обонятельные рецепторы. В полости носа открываются воздухоносные пазухи костей. |
|
| Гортань | Непарные и парные хрящи. Между щитовидным и черпаловидными хрящами натянуты голосовые связки, образующие голосовую щель. Надгортанник прикреплён к щитовидному хрящу. Полость гортани выстлана слизистой оболочкой, покрытой мерцательным эпителием. |
|
| Трахея и бронхи | Трубка 10–13 см с хрящевыми полукольцами. Задняя стенка эластичная, граничит с пищеводом. В нижней части трахея разветвляется на два главных бронха. Изнутри трахея и бронхи выстланы слизистой оболочкой. | Обеспечивает свободное поступление воздуха в альвеолы лёгких. |
| Зона газообмена | ||
| Лёгкие | Парный орган — правое и левое. Мелкие бронхи, бронхиолы, легочные пузырьки (альвеолы).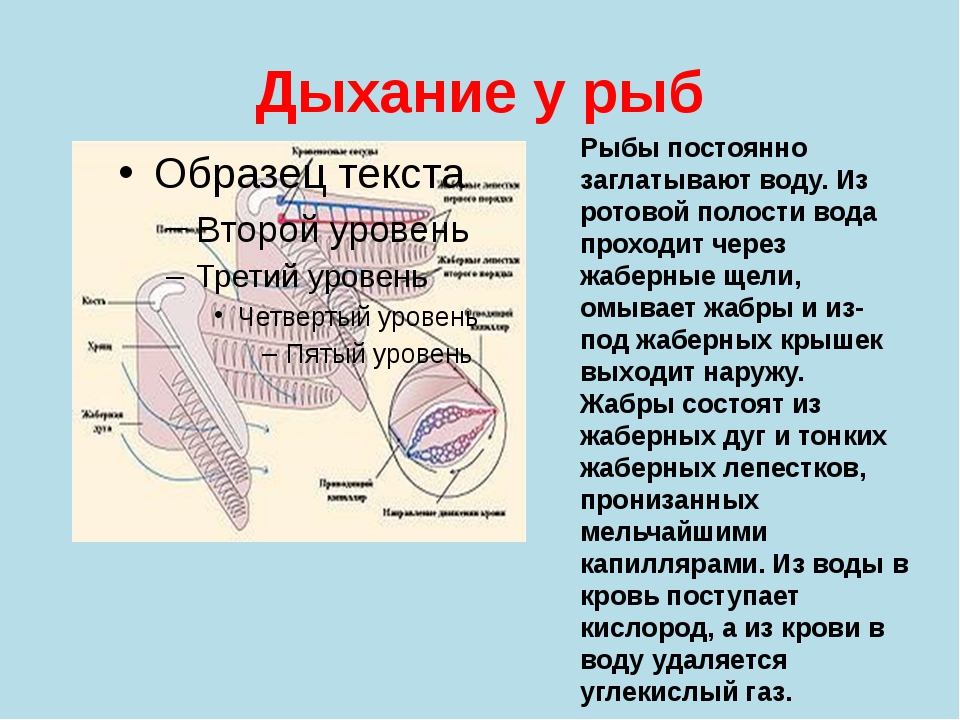 Стенки альвеол образованы однослойным эпителием и оплетены густой сетью капилляров. Стенки альвеол образованы однослойным эпителием и оплетены густой сетью капилляров. |
Газообмен через альвеолярно-капилярную мембрану. |
| Плевра | Снаружи каждое лёгкое покрыто двумя листками соединительнотканной оболочки: легочная плевра прилегает к лёгким, пристеночная — к грудной полости. Между двумя листками плевры — полость (щель), заполненная плевральная жидкостью. |
|

Функции дыхательной системы
- Обеспечение клеток организма кислородом О2.
- Удаление из организма углекислого газа СО2, а также некоторых конечных продуктов обмена веществ (паров воды, аммиака, сероводорода).
Носовая полость
Воздухоносные пути начинаются с носовой полости, которая через ноздри соединяется с окружающей средой. От ноздрей воздух проходит по носовым ходам, выстланным слизистым, реснитчатым и чувствительным эпителием. Наружный нос состоит из костных и хрящевых образований и имеет форму неправильной пирамиды, которая изменяется в зависимости от особенностей строения человека. В состав костного скелета наружного носа входят носовые косточки и носовая часть лобной кости. Хрящевой скелет является продолжением костного скелета и состоит из гиалиновых хрящей различной формы. Полость носа имеет нижнюю, верхнюю и две боковые стенки. Нижняя стенка образована твёрдым нёбом, верхняя — решётчатой пластинкой решётчатой кости, боковая — верхней челюстью, слёзной костью, глазничной пластинкой решётчатой кости, нёбной костью и клиновидной костью. Носовой перегородкой полость носа разделена на правую и левую части. Перегородка носа образована сошником, перпендикулярной пластинкой решётчатой кости и спереди дополняется четырёхугольным хрящом носовой перегородки.
От ноздрей воздух проходит по носовым ходам, выстланным слизистым, реснитчатым и чувствительным эпителием. Наружный нос состоит из костных и хрящевых образований и имеет форму неправильной пирамиды, которая изменяется в зависимости от особенностей строения человека. В состав костного скелета наружного носа входят носовые косточки и носовая часть лобной кости. Хрящевой скелет является продолжением костного скелета и состоит из гиалиновых хрящей различной формы. Полость носа имеет нижнюю, верхнюю и две боковые стенки. Нижняя стенка образована твёрдым нёбом, верхняя — решётчатой пластинкой решётчатой кости, боковая — верхней челюстью, слёзной костью, глазничной пластинкой решётчатой кости, нёбной костью и клиновидной костью. Носовой перегородкой полость носа разделена на правую и левую части. Перегородка носа образована сошником, перпендикулярной пластинкой решётчатой кости и спереди дополняется четырёхугольным хрящом носовой перегородки.
На боковых стенках полости носа располагаются носовые раковины — по три с каждой стороны, что увеличивает внутреннюю поверхность носа, с которой соприкасается вдыхаемый воздух.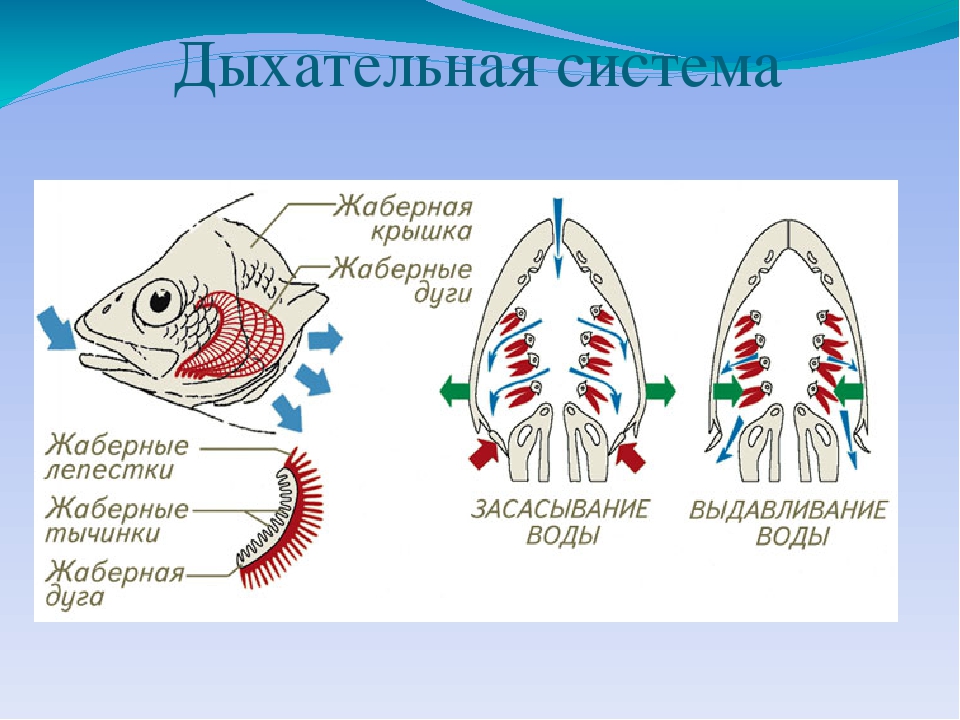
Носовая полость образована двумя узкими и извилистыми носовыми ходами. Здесь воздух согревается, увлажняется и освобождается от частичек пыли и микробов. Оболочка, выстилающая носовые ходы, состоит из клеток, которые выделяют слизь, и клеток реснитчатого эпителия. Движением ресничек слизь вместе с пылью и микробами направляется из носовых ходов наружу.
Внутренняя поверхность носовых ходов богато снабжена кровеносными сосудами. Вдыхаемый воздух, попадает в полость носа, обогревается, увлажняется, очищается от пыли и частично обезвреживается. Из носовой полости он попадает в носоглотку. Затем воздух из носовой полости попадает в глотку, а из неё — в гортань.
Гортань
Гортань — один из отделов воздухоносных путей. Сюда из носовых ходов через глотку поступает воздух. В стенке гортани есть несколько хрящей: щитовидный, черпаловидный и др. В момент глотания пищи мышцы шеи поднимают гортань, а надгортанный хрящ опускается и закрывается гортань. Поэтому пища поступает только в пищевод и не попадает в трахею.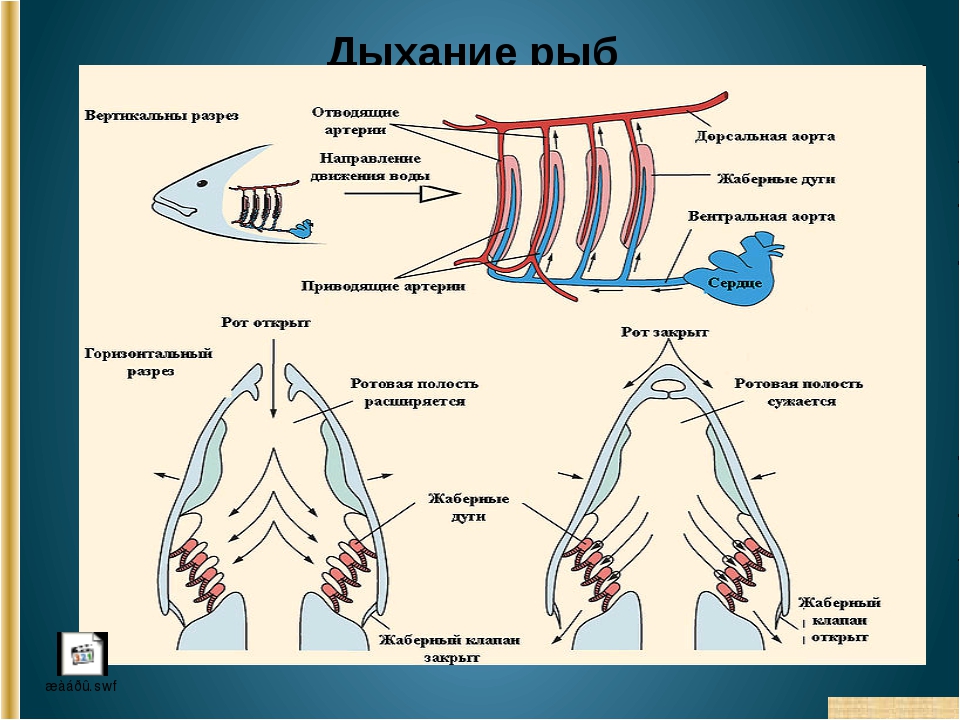
В узкой части гортани расположены голосовые связки, посредине между ними находится голосовая щель. При прохождении воздуха голосовые связки вибрируют, производя звук. Образование звука происходит на выдохе при управляемом человеком движении воздуха. В формировании речи участвуют: носовая полость, губы, язык, мягкое нёбо, мимические мышцы.
Трахея
Гортань переходит в трахею (дыхательное горло), которая имеет форму трубки длиной около 12 см, в стенках которого есть хрящевые полукольца, не позволяющие ей спадать. Задняя стенка её образована соединительнотканной перепонкой. Полость трахеи, как и полость других воздухоносных путей выстлана мерцательным эпителием, препятствующим проникновению в лёгкие пыли и других инородных тел. Трахея занимает серединное положение, сзади она прилежит к пищеводу, а по бокам от неё располагаются сосудисто-нервыне пучки. Спереди шейный отдел трахеи прикрывают мышцы, а вверху она охватывается ещё щитовидной железой. Грудной отдел трахеи прикрыт спереди рукояткой грудины, остатками вилочковой железы и сосудами. Изнутри трахея покрыта слизистой оболочкой, содержащей большое количество лимфоидной ткани и слизистых желёз. При дыхании мелкие частички пыли прилипают к увлажнённой слизистой оболочке трахеи, а реснички мерцательного эпителия продвигают их обратно к выходу из дыхательных путей.
Нижний конец трахеи делится на два бронха, которые затем многократно ветвятся, входят в правое и левое лёгкие, образуя в лёгких «бронхиальное дерево».
Бронхи
В грудной полости трахея делится на два бронха — левый и правый. Каждый бронх входит в лёгкое и там делится на бронхи меньшего диаметра, которые разветвляются на мельчайшие воздухоносные трубочки — бронхиолы. Бронхиолы в результате дальнейшего ветвления переходят в расширения — альвеолярные ходы, на стенках которых находятся микроскопические выпячивания, называемые легочными пузырьками, или альвеолами.
Стенки альвеол построены из особого тонкого однослойного эпителия и густо оплетены капиллярами. Общая толщина стенки альвеолы и стенки капилляра составляет 0,004 мм. Через эту тончайшую стенку происходит газообмен: в кровь из альвеолы поступает кислород, а обратно — углекислый газ. В лёгких насчитывается несколько сотен миллионов альвеол. Общая поверхность их у взрослого человека составляет 60–150 м2. благодаря этому в кровь поступает достаточное количество кислорода (до 500 литров в сутки).
Лёгкие
Лёгкие занимают почти всю полость грудной полости и представляют собой упругие губчатые органы. В центральной части лёгкого располагаются ворота, куда входят бронх, легочная артерия, нервы, а выходят легочные вены. Правое лёгкое делится бороздами на три доли, левое на две. Снаружи лёгкие покрыты тонкой соединительнотканной плёнкой — легочной плеврой, которая переходит на внутреннею поверхность стенки грудной полости и образует пристенную плевру. Между этими двумя плёнками находится плевральная щель, заполненная жидкостью, уменьшающей трение при дыхании.
На лёгком различают три поверхности: наружную, или рёберную, медиальную, обращённую в сторону другого лёгкого, и нижнюю, или диафрагмальную. Кроме того, в каждом лёгком различают два края: передний и нижний, отделяющие диафрагмальную и медиальную поверхности от рёберной. Сзади рёберная поверхность без резкой границы переходит в медиальную. Передний край левого лёгкого имеет сердечную вырезку. На медиальной поверхности лёгкого располагаются его ворота. В ворота каждого лёгкого входит главный бронх, легочная артерия, которая несёт в лёгкое венозную кровь, и нервы, иннервирующие лёгкое. Из ворот каждого лёгкого выходят две легочные вены, которые несут к сердцу артериальную кровь, и лимфатические сосуды.
Лёгкие имеют глубокие борозды, разделяющие их на доли — верхнюю, среднюю и нижнюю, а в левом две — верхнюю и нижнюю. Размеры лёгкого не одинаковы. Правое лёгкое несколько больше левого, при этом оно короче его и шире, что соответствует более высокому стоянию правого купола диафрагмы в связи с правосторонним расположением печени. Цвет нормальных лёгких в детском возрасте бледно-розовый, а у взрослых они приобретают тёмно-серую окраску с синеватым оттенком — следствие отложения в них попадающих с воздухом пылевых частиц. Ткань лёгкого мягкая, нежная и пористая.
Газообмен лёгких
В сложном процессе газообмена выделяют три основные фазы: внешнее дыхание, перенос газа кровью и внутреннее, или тканевое, дыхание. Внешнее дыхание объединяет все процессы, происходящие в лёгком. Оно осуществляется дыхательным аппаратом, к которому относятся грудная клетка с мышцами, приводящими её в движение, диафрагма и лёгкие с воздухоносными путями.
Воздух, поступивший в лёгкие при вдохе, изменяет свой состав. Воздух в лёгких отдаёт часть кислорода и обогащается углекислым газом. Содержание углекислого газа в венозной крови выше, чем в воздухе, находящемся в альвеолах. Поэтому углекислый газ выходит из крови в альвеолы и содержание его меньше, чем в воздухе. Сначала кислород растворяется в плазме крови, далее связывается с гемоглобином, а в плазму поступают новые порции кислорода.
Переход кислорода и углекислого газа из одной среды в другую проходит благодаря диффузии от большей концентрации к меньшей. Хотя диффузия протекает медленно, поверхность контакта крови с воздухом в лёгких настолько велика, что полностью обеспечивает нужный газообмен. Подсчитано, что полный газообмен между кровью и альвеолярным воздухом может происходить за время, которое втрое короче, чем время пребывания крови в капиллярах (т.е. в организме имеются значительные резервы обеспечения тканей кислородом).
Венозная кровь, попав в лёгкие, отдаёт углекислый газ, обогащается кислородом и превращается в артериальную. В большом круге эта кровь расходится по капиллярам во все ткани и отдаёт кислород клеткам тела, которые постоянно потребляют его. Углекислого газа, выделяющегося клетками в результате их жизнедеятельности, здесь больше, чем в крови, и он диффундирует из тканей в кровь. Таким образом, артериальная кровь, пройдя через капилляры большого круга кровообращения, становится венозной и правой половиной сердца направляется в лёгкие, здесь опять насыщается кислородом и отдаёт углекислый газ.
В организме дыхание осуществляется с помощью дополнительных механизмов. Жидкие среды, входящие в состав крови (её плазмы), обладают низкой растворимостью в них газов. Поэтому, для того чтобы человек мог существовать, ему нужно было бы иметь сердце мощнее в 25 раз, лёгкие — в 20 раз и за одну минуту перекачивать более 100 литров жидкости (а не пять литров крови). Природа нашла способ преодоления этой трудности, приспособив для переноса кислорода особое вещество — гемоглобин. Благодаря гемоглобину кровь способна связывать кислород в 70 раз, а углекислый газ — в 20 раз больше, чем жидкая часть крови — её плазма.
Альвеола — тонкостенный пузырёк диаметром 0,2 мм, заполненный воздухом. Стенка альвеолы образована одним слоем плоских клеток эпителия, по наружной поверхности которых разветвляется сетка капилляров. Таким образом, газообмен происходит через очень тонкую перегородку, образованную двумя слоями клеток: стенки капилляра и стенки альвеолы.
Обмен газов в тканях (тканевое дыхание)
Обмен газов в тканях осуществляется в капиллярах по тому же принципу, что и в лёгких. Кислород из тканевых капилляров, где его концентрация высока, переходит в тканевую жидкость с более низкой концентрацией кислорода. Из тканевой жидкости он проникает в клетки и сразу же вступает в реакции окисления, поэтому в клетках практически нет свободного кислорода.
Диоксид углерода по тем же законам поступает из клеток, через тканевую жидкость, в капилляры. Выделяющийся углекислый газ способствует диссоциации оксигемоглобина и сам вступает в соединение с гемоглобином, образуя карбоксигемоглобин, транспортируется в лёгкие и выделяется в атмосферу. В оттекающей от органов венозной крови углекислый газ находится как в связанном, так и в растворённом состоянии в виде угольной кислоты, которая в капиллярах лёгких легко распадается на воду и углекислый газ. Угольная кислота может также вступать в соединения с солями плазмы, образуя бикарбонаты.
В лёгких, куда поступает венозная кровь, кислород снова насыщает кровь, а углекислый газ из зоны высокой концентрации (легочных капилляров) переходит в зону низкой концентрации (альвеол). Для нормального газообмена воздух в лёгких постоянно сменяться, что достигается ритмическими атаками вдоха и выдоха, за счёт движений межрёберных мышц и диафрагмы.
Транспорт кислорода в организме
| Путь кислорода | Функции |
| Верхние дыхательные пути | |
| Носовая полость | Увлажнение, согревание, обеззараживание воздуха, удаление частиц пыли |
| Глотка | Проведение согретого и очищенного воздуха в гортань |
| Гортань | Проведение воздуха из глотки в трахею. Защита дыхательных путей от попадания пищи надгортанным хрящом. Образование звуков путём колебания голосовых связок, движения языка, губ, челюсти |
| Трахея | Свободное продвижение воздуха |
| Бронхи | Свободное продвижение воздуха |
| Лёгкие | Органы дыхания. Дыхательные движения осуществляются под контролем центральной нервной системы и гуморального фактора, содержащегося в крови, — СО2 |
| Альвеолы | Увеличивают площадь дыхательной поверхности, осуществляют газообмен между кровью и лёгкими |
| Кровеносная система | |
| Капилляры лёгких | Транспортируют венозную кровь из легочной артерии в лёгкие. По законам диффузии О2 поступает из мест большей концентрации (альвеолы) в места меньшей концентрации (капилляры), в то же время СО2 диффундирует в противоположном направлении. |
| Легочная вена | Транспортирует О2 от лёгких к сердцу. Кислород, попав в кровь, сначала растворяется в плазме, затем соединяется с гемоглобином, и кровь становится артериальной |
| Сердце | Проталкивает артериальную кровь по большому кругу кровообращения |
| Артерии | Обогащают кислородом все органы и ткани. Легочные артерии несут венозную кровь к лёгким |
| Капилляры тела | Осуществляют газообмен между кровью и тканевой жидкостью. О2 переходит в тканевую жидкость, а СО2 диффундирует в кровь. Кровь становится венозной |
| Клетка | |
| Митохондрии | Клеточное дыхание — усвоение О2 воздуха. Органические вещества благодаря О2 и дыхательным ферментам окисляются (диссимиляция) конечные продукты — Н2О, СО2 и энергия которая идёт на синтез АТФ. Н2О и СО2 выделяются в тканевую жидкость, из которой диффундируют в кровь. |
Значение дыхания.
Дыхание — это совокупность физиологических процессов, обеспечивающих газообмен между организмом и внешней средой (внешнее дыхание), и окислительных процессов в клетках, в результате которых выделяется энергия (внутреннее дыхание). Обмен газов между кровью и атмосферным воздухом (газообмен) — осуществляется органами дыхания.
Источником энергии в организме служат пищевые вещества. Основным процессом, освобождающим энергию этих веществ, является процесс окисления. Он сопровождается связыванием кислорода и образованием углекислого газа. Учитывая, что в организме человека нет запасов кислорода, непрерывное поступление его жизненно необходимо. Прекращение доступа кислорода в клетки организма ведёт к их гибели. С другой стороны, образованный в процессе окисления веществ углекислый газ должен быть удалён из организма, так как накопление значительного количества его опасно для жизни. Поглощение кислорода из воздуха и выделение углекислого газа осуществляется через систему органов дыхания.
Биологическое значение дыхания заключается в:
- обеспечении организма кислородом;
- удалении углекислого газа из организма;
- окислении органических соединений БЖУ с выделением энергии, необходимой человеку для жизнедеятельности;
- удалении конечных продуктов обмена веществ (пары воды, аммиака, сероводорода и т.д.).
|
ЗНАЕТЕ ЛИ ВЫ?
|
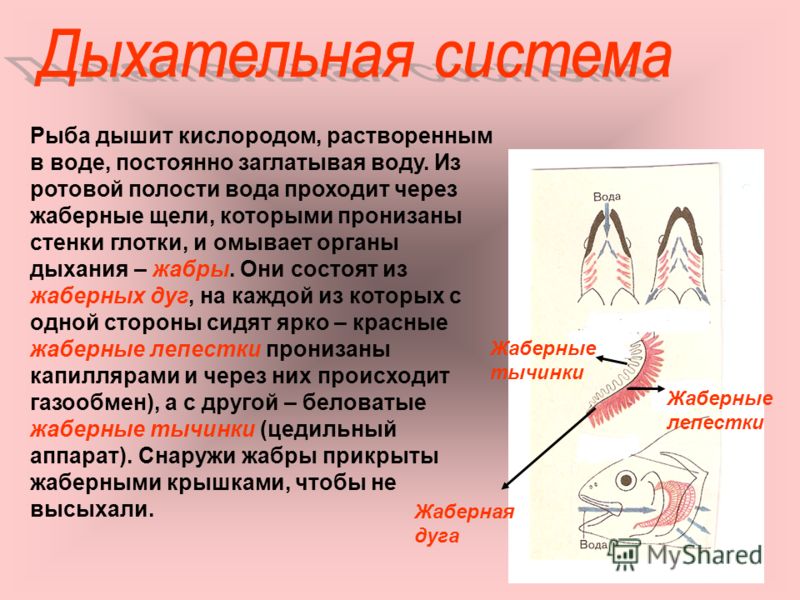
Большинство рыб дышат растворённым в воде кислородом, но есть виды, приспособившиеся частично и к воздушному дыханию (двоякодышащие, прыгун, змееголов и др.).
Форма жабр разнообразна и зависит от вида рыб и их подвижности:
В табл. 16. представлены основные органы извлечения кислорода из воды в разные возрастные периоды
Таблица 16
Органы дыхания рыб в разные возрастные периоды
Таблица 17
Площадь жаберной поверхности у некоторых видов рыб
Рис. 18. Органы дыхания у эмбрионов рыб: а – пелагическая рыба; б – карп; в – вьюн; 1 – Кювьеровы протоки; 2 – нижняя хвостовая вена; 3 – сеть капилляров; 4 – наружные жабры
Рис. 19. Схема противотока крови и воды в жабрах рыб: 1 – хрящевой стержень; 2 – жаберная дуга; 3 – жаберные лепестки; 4 – жаберные пластинки; 5 – приносящая артерия от брюшной аорты; 6 – выносящая артерия к спинной аорте
Рис. 20. Механизм дыхания взрослой рыбы: а – вдох; б – выдох
Читайте также:
|
Система дыхания у рыб | Fishong
Чем же дышат рыбы? Конечно, кислородом! Только тем, который растворяется в водоеме. Их органы дыхания – это жабры, которые состоят из огромного количества лепестков, имеющих кровеносные сосуды. Огромную поверхность имеют жаберные пластинки. К примеру, у окуня поверхность тела в тридцать раз меньше ее.
Если вы внимательно когда-нибудь следили за рыбой, то могли отметить, что она имеет привычку открывать и закрывать рот. Такой же процесс происходит с рыбьими жабрами, когда они открываются, то закрывается рот, дальше идет все наоборот.
Так у рыбы происходит заглатывание воды, после чего она закрывает рот, дальше вода перебирается в жабры и через щель у них протекает наружу. И в этом процессе именно кровеносная система жабр поглощает кислород, таким образом, обогащая им кровь.
Для каждого рыбного вида есть свой определенный порог кислорода, который содержится в воде. Когда такой порог понижается, то рыбные особи могут становиться неактивными и очень вялыми, а затем, и вовсе умирают. Такую рыбу еще называют замора.
Есть и такая рыба, которая тогда, когда в воде присутствует мало кислорода, может заглатывать атмосферный воздух – это крась и другие особи. Если взять, к примеру, окуня, то в его функцию дыхания может включаться плавательный пузырь, который пронизывается капиллярными сосудами. Дополнительное дыхание имеет и сом – это кожное.
Обогащается водоем кислородом посредством атмосферного воздуха. А на сам этот процесс влияет множество факторов:
- температурный режим воды;
- какого объема водоем;
- есть ли подземные родники или ключи;
- сколько слоев воды примешивается.
Если летом температура несколько снизывается, а также стоит ветряная погода, то это способствует тому, что кислород очень хорошо растворяется в воде. Вот в такой период рыба себя очень хорошо чувствует даже в таких водоемах, где до этого кислорода было недостаточно.
Когда рыба оживает и ее самочувствие улучшается – это означает, что и клев будет хорошим. Так что рыболову обязательно нужно поинтересоваться погодными условиями на момент его рыбалки.
Уже не одним опытным рыбаком замечено, что после дождевой погоды активность рыб повышается а, значит, клев рыбы будет усиленным. И это понятно, ведь капельки дождя хорошо насыщены кислородом, а это, в свою очередь, увеличивает его содержание в водоемах, где в это время воздух перемешивается с водой.
В холодное, зимнее время рыбы ищут глубокие места, где дно может быть, как песчаное, так и каменистое. А также те места, где происходит впадение речек или ручьев или там, где выходят ключевые воды.
Так что рыбам, как и другим существам, кислород жизненно необходим. Только дыхательные органы у них устроены по-другому. Вы никогда не задумывались над такой аномалией, почему, если рыбе в воде не хватает кислорода, то вытащив ее из водоема, она умирает от кислородного голодания. А ведь воздух насыщен кислородом намного больше, чем вода. Но вся причина заключается в жабрах, вне воды их ламеллы начинают разрушаться. Рыбам нельзя находиться дольше, чем две минуты вне водного пространства.
Дыхательная система рыб. Специфические особенности строения рыб
Благодаря тому, что каждое существо наделено органами дыхания, все мы получаем то, без чего не можем жить – кислород. У всех наземных животных и людей эти органы называются легкими, которые поглощают максимальное количество кислорода из воздуха. Дыхательная система рыб же состоит из жабр, которые втягивают в организм кислород из воды, где его куда меньше, чем в воздухе. Именно из-за этого строение тела данного биологического вида так отличается от всех хребетных наземных существ. Что же, рассмотрим все особенности строения рыб, их дыхательной системы и прочих жизненно важных органов.
Кратко о рыбах
Для начала попробуем разобраться в том, что же это за существа, как и чем они живут, какую имеют взаимосвязь с человеком. Потому сейчас мы начинаем наш урок биологии, тема “Морские рыбы”. Это надкласс позвоночных животных, которые обитают исключительно в водной среде. Характерной чертой является то, что все рыбы челюсторотные, а также обладают жабрами. Отметить стоит, что данные показатели характерны для каждого вида рыб, вне зависимости от размера и массы. В жизни человека данный подкласс играет экономически важную роль, так как большинство его представителей употребляются в пищу.
Считается также, что рыбы были на заре эволюции. Именно такие существа, которые могли обитать под водой, но еще не имели челюстей, когда-то были единственными жителями Земли. С тех пор вид эволюционировал, некоторые из них превратились в животных, некоторые остались под водой. Вот и весь урок биологии. Тема “Морские рыбы. Краткий экскурс в историю” рассмотрена. Наука, изучающая морские рыбы, носит название “ихтиология”. Давайте теперь перейдем к изучению этих существ с более профессиональной точки зрения.
Общая схема строения рыб
Обобщенно можно сказать, что тело каждой рыбы делится на три части – голова, туловище и хвост. Голова заканчивается в районе жабр (в их начале или конце – зависит от надкласса). Туловище оканчивается на линии анального отверстия у всех представителей данного класса морских обитателей. Хвост же – простейшая часть организма, которая состоит из стержня и плавника.
Форма тела строго зависит от условий обитания. Рыба, которая живет в средней толще воды (лосось, акула), имеет торпедовидную фигуру, реже – стреловидную. Те же морские обитатели, которые плавают над самым дном, имеют сплющенную форму. К ним можно отнести камбалу, морских лисиц и других рыбок, которые вынуждены плавать среди растений или камней. Они приобретают более маневренные очертания, которые имеют много общего со змеями. К примеру, угорь является обладателем сильно вытянутого тела.
Визитка рыбы – ее плавники
Без плавников невозможно себе представить строение рыбы. Картинки, которые представлены даже в детских книгах, непременно демонстрируют нам эту часть тела морских жителей. Что же они собой представляют?
Итак, плавники бывают парными и непарными. К парным можно отнести грудные и брюшные, которые симметричны и синхронно двигаются. Непарные представлены в виде хвоста, спинных плавников (от одного до трех), а также анального и жирового, который находится сразу сзади спинного. Сами по себе плавники состоят из жестких и мягких лучей. Именно исходя из количества этих лучей высчитывается плавниковая формула, которая применяется для определения конкретного вида рыбы. Латинскими буквами определяется местоположение плавника (А – анальный, P – грудной, V – брюшной). Далее римскими цифрами указывается количество жестких лучей, а арабскими – мягких.
Классификация рыб
Сегодня условно всех рыб можно разделить на две категории – хрящевые и костные. В первую группу входят такие обитатели моря, скелет которых состоит их хрящей различного размера. Это вовсе не означает, что подобное существо мягкое и не способное к передвижению. У многих представителей надкласса хрящи затвердевают, и по своей плотности становятся почти как кости. Вторая категория – костные рыбы. Биология как наука утверждает, что именно этот надкласс был отправной точкой эволюции. Некогда в его рамках находилась давно вымершая кистеперая рыба, от которой, возможно, произошли все наземные млекопитающие. Далее мы более подробно рассмотрим строение тела рыбы каждого из этих видов.
Хрящевые
В принципе, строение хрящевых рыб не являет собой нечто сложно и необычное. Это обыкновенный скелет, который состоит из очень твердых и прочных хрящей. Каждое соединение пропитано солями кальция, благодаря которым в хрящах и появляется прочность. Хорда держит свою форму на протяжении всей жизни, при этом она частично редуцирована. Череп соединен с челюстями, вследствие чего скелет рыбы имеет целостную структуру. К нему также присоединены плавники – хвостовой, парные брюшные и грудные. Челюсти располагаются на брюшной стороне скелета, а над ними находятся две ноздри. Хрящевой скелет и мышечный корсет таких рыб снаружи покрыт плотной чешуей, которая называется плакоидной. Она состоит из дентина, который по своему составу похож на обыкновенные зубы у всех наземных млекопитающих.
Как хрящевые дышат
Дыхательная система рыб надкласса хрящевых представлена в первую очередь жаберными щелями. Их насчитывают от 5 до 7 пар на теле. Во внутренние органы кислород распространяется благодаря спиральному клапану, который тянется вдоль всего организма рыбы. Характерной чертой всех хрящевых является то, что у них отсутствует плавательный пузырь. Именно поэтому они вынуждены постоянно находиться в движении, чтобы не пойти ко дну. Важно также отметить, что в организме хрящевых рыб, которые априори обитают в соленых водах, содержится минимальное количество этой самой соли. Ученые полагают что это связано с тем, что в крови у данного надкласса очень много мочевины, которая состоит преимущественно из азота.
Костные
Теперь рассмотрим, как выглядит скелет рыбы, принадлежащий к надклассу костных, а также узнаем, чем еще характерны представители этой категории.
Итак, скелет представлен в виде головы, туловища (они существуют отдельно, в отличие от предыдущего случая), а также парных и непарных конечностей. Черепная коробка поделена на два отдела – мозговой и висцеральный. Второй включает в себя челюстную и подъязычную дуги, которые являются главными составляющими челюстного аппарата. Также в скелете костной рыбы имеются жаберные дуги, которые предназначены для удержания жаберного аппарата. Что касается мышц данного вида рыб, то все они имеют сегментарное строение, а наиболее развитые из них – это челюстные, плавниковые и жаберные.
Дыхательный аппарат костных обитателей моря
Наверное, уже стало всем понятно, что дыхательная система рыб надкласса костных главным образом состоит из жабр. Они располагаются на жаберных дугах. Также неотъемлемой составляющей частью таких рыб являются жаберные щели. Они прикрыты одноименной крышкой, которая предназначена для того, чтобы рыба могла дышать даже в обездвиженном состоянии (в отличие от хрящевых). Некоторые представители надкласса костных могут дышать через кожный покров. А вот те, которые обитают непосредственно под поверхностью воды, и при этом никогда глубоко не опускаются, наоборот, захватывают воздух своими жабрами из атмосферы, а не из водной среды.
Строение жабр
Жабры – уникальный орган, который ранее был присущ всем первичноводным созданиям, проживавшим на Земле. В нем происходит процесс газообмена между гидросредой и организмом, в котором они функционируют. Жабры рыбы нашего времени мало чем отличаются от тех жабр, которые были присущи более ранним обитателям нашей планеты.
Как правило, они представлены в виде двух одинаковых пластинок, которые пронизаны весьма густой сетью кровеносных сосудов. Неотъемлемой частью жабр является целомическая жидкость. Именно она совершает процесс газообмена между водной средой и организмом рыбы. Отметим, что данное описание дыхательной системы присуще не только рыбам, а многим позвоночным и не позвоночным обитателям морей и океанов. А вот о том, что особенного в себе несут именно те органы дыхания, которые находятся в организме рыб, читайте далее.
Где располагаются жабры
Дыхательная система рыб в своем большинстве сосредоточена в глотке. Именно там располагаются жаберные дуги, на которых закреплены одноименные органы газообмена. Они представлены в виде лепестков, которые пропускают сквозь себя и воздух, и различные жизненно-необходимые жидкости, что находятся внутри каждой рыбы. В определенных местах глотка пронизывается жаберными щелями. Именно через них проходит тот кислород, который поступает в рот рыбы с заглатываемою ею водой.
Весьма важным фактом является то, что в сравнении с размерами организма многих морских обитателей, их жабры весьма велики для них. В связи с этим в их организмах возникают проблемы с осмолярностью плазмы крови. Из-за этого рыбы всегда пьют морскую воду и выпускают ее через жаберные щели, тем самым ускоряя различные обменные процессы. Она имеет меньшую консистенцию, нежели кровь, потому быстрее и эффективнее снабжает жабры и прочие внутренние органы кислородом.
Сам процесс дыхания
Когда рыба только появляется на свет, дышит практически все ее тело. Кровеносными сосудами пронизан каждый ее орган, включая наружную оболочку, потому кислород, который находится в морской воде, проникает в организм постоянно. Со временем у каждой подобной особи начинает развиваться жаберное дыхание, так как наибольшей сеткой кровеносных сосудов оснащаются именно жабры и все прилегающие к ним органы. Тут то и начинается самое интересное. Процесс дыхания каждой рыбы зависит от ее анатомических особенностей, потому в ихтиологии принято делить его на две категории – активное дыхание и пассивное. Если с активным все понятно (рыба дышит «обычно», набирая кислород в жабры и обрабатывая его, как человек), то с пассивным мы сейчас попробуем разобраться более детально.
Пассивное дыхание и от чего оно зависит
Данный тип дыхания свойственен только быстроходным обитателям морей и океанов. Как мы уже говорили выше, акулы, а также некоторые другие представители хрящевого надкласса не могут длительное время находиться без движения, так как у них отсутствует плавательный пузырь. Этому есть еще одна причина, а именно – это и есть пассивное дыхание. Когда рыба плывет на большой скорости, она приоткрывает рот, и туда автоматически попадает вода. Приближаясь к трахеям и жабрам, от жидкости отделяется кислород, который и питает организм морского скороходного обитателя. Именно поэтому длительное время находясь без движения рыба лишает себя возможности дышать, не затрачивая на это никаких сил и энергии. Напоследок заметим, что к таким быстроходным жителям соленых вод относятся преимущественно акулы и все представители скумбриевых.
Главная мышца организма рыбы
Весьма простым является строение сердца рыбы, которое, отметим, за всю историю существования данного класса животных, практически не эволюционировало. Итак, этот орган у них двухкамерный. Он представлен одним основным насосом, в состав которого входит две камеры – предсердие и желудочек. Рыбье сердце перекачивает только венозную кровь. В принципе, система кровообращения у данного вида морских обитателей имеет замкнутую систему. Кровь циркулирует через все капилярчики жабр, затем сливается в сосудах, а оттуда снова расходится на более мелкие капилляры, которые уже снабжают остальные внутренние органы. После этого «отработанная» кровь собирается в венах (их у рыб две – печеночная и кардиальная), откуда уже поступает непосредственно к сердцу.
Заключение
Вот и подошел к концу наш краткий урок биологии. Тема рыб, как оказалось, весьма интересна, увлекательна и проста. Организм данных обитателей моря крайне важен для изучения, так как считается, что именно они были первыми обитателями нашей планеты, каждая из них – это есть ключ к разгадке эволюции. Кроме того, изучать строение и функционирование рыбьего организма намного проще, чем какого-либо другого. И размеры данных обитателей водной стохии вполне приемлемы для детального рассмотрения, и при этом все системы и образования просты и доступны даже для детей школьного возраста.
хрящевые и костные 🐲 СПАДИЛО.РУ
Рыбы — надкласс позвоночных животных, включает в себя два класса: хрящевые и костные.
Класс Хрящевые рыбы (Hondrichthyes)
К этому классу относят около 730 видов. В процессе эволюции хрящевые появились раньше костных рыб. Их скелет состоит из хряща и не окостеневает до конца жизни. Класс разделяют на пластинчатожаберных и химеровых. рассмотрим строение плстинчатожаберных рыб на примере типичного представителя – акулы.
Тело по форме напоминает торпеду, сплющено в спинно-брюшном направлении. У донных рыб уплощение выражено сильнее. На нижней стороне головы находится рот, кзади от него по бокам находятся 5 пар жаберных щелей. По бокам тела располагаются парные грудные и брюшные плавники, на спине – два непарных плавника. Может также иметься анальный плавник.
Хвост ассиметричный, верхняя лопасть длиннее и шире, потому что в неё продолжается позвоночный столб. Такое строение хвостового плавника называется гетероцеркальным. У костных рыб встречаются другие типы строения (см. рисунок).
Хвостовой плавник является основным движителем рыб, остальные плавники служат для равновесия или изменения направления.
Тело хрящевых покрыто плакоидной чешуёй. Она состоит из множества пластинок с заострёнными зубцами и поэтому похожа на наждачную бумагу. Чешуйки, перешедшие на челюсть, становятся зубами, поэтому у акул может быть 5-10 рядов зубов.
Скелет
Осевой скелет образован позвоночным столбом и черепом. Позвоночник приходит на смену хорде в эмбриональном периоде. Между верхней и нижней дугами позвонков проходит спинной мозг. В туловищном отделе к позвонкам прикрепляются короткие рёбра.
Череп разделён на два отдела: мозговой и висцеральный. Висцеральный череп включает в себя челюсть, подъязычную дугу и жаберные дуги, на которых расположены жабры.
Конечности рыб – плавники. Парные плавники прикреплены к дугообразным хрящам, лежащим в мышцах тела, и образуют передний и задний пояса конечностей.
Пищеварительная система
Так как зубы хрящевых рыб образуются из чешуи, они сменяются в течении жизни. За несколько лет акула может сменить до 20 000 зубов.
Ротовое отверстие имеет вид косой щели. За ротовой полостью в пищеварительном тракте следует глотка, в которую открываются жаберные щели. Некоторые акулы фильтруют воду жаберными щелями, чтобы добыть питательный планктон, однако большинство видов хищники или падальщики.
Короткий пищевод ведёт из глотки в желудок, где пища обрабатывается ферментами (в основном, пепсином). Содержимое желудка имеет кислую реакцию. Через желудочный сфинктер частично переработанная пища попадает в тонкую кишку. Туда же открываются протоки поджелудочной железы и печени. В тонкой кишке создаётся щелочная среда.
Толстая кишка короткая, но имеет винтообразную складку стенки – спиральный клапан, что увеличивает площадь поверхности. Толстая кишка переходит в прямую, та открывается в клоаку. Также в клоаку открываются протоки половых желёз и мочеточника.
Дыхательная система
Жаберный аппарат расположен на хрящевых дугах. В просвет щелей выступает множество выростов – жаберных лепестков. Газообмен происходит при движении воды из глотки наружу, поэтому для дыхания акулы вынуждены постоянно двигаться или активно засасывать воду.
Кровеносная система
Кровь рыб красного цвета, содержит форменные элементы, в эритроцитах есть ядра.
Сердце состоит из предсердия и желудочка. Венозная кровь с низким содержанием кислорода из желудочка сердца попадает в единственный круг кровообращения. По брюшной аорте она идёт в сторону головы, к жабрам. В жабрах кровь насыщается кислородом, собирается в спинную аорту и разносится по всему телу, насыщая ткани и органы.
От органов пищеварения кровь направляется по воротной вене в печень, где очищается от токсических веществ и избавляется от избытка моносахаридов.
В хвостовой части происходит очищение от продуктов обмена: по хвостовой вене кровь попадает в почки.
Также у рыб имеется лимфатическая система, состоящая из тонкостенных сосудов, лимфатических узлов нет.
Выделительная система
Рыбы обладают туловищными почками (мезонефросами). Внешне они напоминают метанефридии кольчатых червей, но устроены сложнее.
В почечном клубочке происходит фильтрация крови, а не полостной жидкости, как у более примитивных животных. Образуется первичная моча, близкая по составу к плазме крови. При дальнейшем прохождении мочи по канальцу полезные для организма продукты всасываются обратно в кровь, а токсичные продукты обмена, наоборот, выводятся. В итоге образуется вторичная моча, которая выводится по мочеточнику. Структурно-функциональная единица почки – нефрон.
Половая система
Мужская половая система хрящевых рыб анатомически объединена с мочевыделительной. Протоки парных семенников впадают в мочеточник, который по совместительству является и семяпроводом.
Женская половая система устроена иначе. Гаметы из яичников сперва попадают в полость тела, а оттуда по яйцеводу идут к клоаке.
Для хрящевых рыб характерно внутренне оплодотворение. Пока оплодотворённая яйцеклетка движется по яйцеводу, она покрывается секретом скорлуповых желёз. В результате из клоаки выходит оформленное яйцо.
Большая часть акул откладывает по одному крупному яйцу, из которого через некоторое время выходит молодое животное. У некоторых хрящевых рыб яйца не покидают тело матери и развитие эмбриона полностью происходит в «матке».
Нервная система
У акул есть спинной и головной мозг, развитая сеть периферических нервов. В мозге различают белое и серое вещество. Так как акулы – подвижные хищники, головной мозг у них развит лучше, чем у других хрящевых. Он состоит из мозжечка, продолговатого, среднего, промежуточного и конечного отделов.
По бокам головы у акул расположены глаза. Так как рыбы не могут активно двигать головой, глаза обладают большой подвижностью. В черепе находится внутреннее ухо, воспринимающее колебания воды. Перед ротовым отверстием расположена пара ноздрей, ведущих в обонятельные мешки.
Орган чувств, который есть только у рыб и водных амфибий – боковая линия. Это полая трубка, которая проходит под кожей вдоль всего тела животного, снаружи в неё ведёт множество каналов. На дне трубки находятся рецепторные клетки. Вода по каналам попадает в трубку и оказывает определённое давление на рецепторы. По изменению этого давления рыба может находить добычу, ориентироваться в полной темноте, определять глубину погружения и т.д.
Костные рыбы (Osteichthyes)
Костные рыбы – самый распространённый класс рыб, к нему относятся около 25 000 видов. Выделяют подклассы хрящекостных, лучепёрых, кистепёрых и двоякодышащих.
Форма тела значительно варьирует в зависимости от образа жизни. Основным органом движения, как и у хрящевых, является хвост. Парные плавники могут видоизменяться, позволяя некоторым рыбам ползать по суше, планировать в воздухе, присасываться к предметам или другим животным.
Также у костных рыб есть непарные спиной и анальный плавники, у некоторых видов имеются добавочные плавники и выросты. Они также могут служить для разных целей: защищать от хищников, нести ядовитые шипы, образовывать парус и т.д.
Тело покрыто кожей с чешуёй. Многочисленные железы выделяют слизь, которая выполняет самые разные функции: защищает рыб от бактерий, принимает участие в водно-солевом обмене, служит для распознавания и сигнализации, может быть токсичной.
Чешуя состоит из костных чешуек, расположенных по принципу черепицы. На протяжении жизни число чешуек особи практически не меняется. Чешуйки растут вместе с рыбой, на них образуются годовые кольца.
Скелет
Не у всех костных рыб скелет окостеневает полностью. Так, у хрящекостных (к ним относятся осетровые) есть хрящевые элементы.
Кости рыб могут развиваться двумя способами: из хряща или образовываться в дерме кожи.
Осевой скелет костных рыб мало отличается от скелета хрящевых. Их ребра длиннее и ограничивают полость тела не только с верху, но и с боков. Череп более развит, кости висцерального отдела имеют смешанное происхождение и соединены неподвижно. В висцеральном отделе присутствуют жаберные крышки, которые прикрывают жаберные щели. Они принимают активное участие в токе воды через жабры.
Пищеварительная система
Состоит из тех же отделов, что у хрящевых. Формы строения ротового аппараты разнообразны и зависят от типа питания рыб. Зубы состоят из дентина, покрытого эмалью, могут прирастать к кости или быть подвижными. Некоторые рыбы не имеют зубов.
Желудок выражен слабо или может отсутствовать (например, у карповых). Тонкая кишка значительно длиннее, чем у хрящевых. Поджелудочной железы нет, её функции выполняет многолопастная печень. Спиральный клапан у высших костных отсутствует.
Кишечник открывается анальным отверстием во внешнюю среду, клоака есть только у двоякодышащих.
Органом рыб, который развился из пищеварительного тракта, является плавательный пузырь. Он может быть обособленным или сообщённым с кишечником. Рыба произвольно меняет количество воздуха в плавательном пузыре и таким образом погружается или всплывает. Хрящевые рыбы, у которых пузыря нет, вынуждены всё время двигаться, чтобы оставаться на нужной глубине.
Дыхательная система
Межжаберные перегородки у большинства видов редуцировались, поэтому жаберные лепестки располагаются прямо на дугах, с обеих сторон. При поднятии жаберных крышек вода засасывается в ротовое отверстие. При закрытии крышек вода выходит через жаберные щели, омывая лепестки.
Таким образом, костные рыбы могут дышать в стоячей воде, находясь на одном месте. В условиях дефицита кислорода возможен также кожный газообмен. У двоякодышащих и некоторых ильных рыб имеется ячеистое лёгкое (видоизменённый плавательный пузырь).
Кровеносная система
Сходна по строению с кровеносной системой хрящевых. Лимфатическая система развита лучше.
Выделительная система
Электролитный состав жидкостей хрящевых рыб близок к морской воде, поэтому поддерживать водно-солевой баланс им легче. Выделительная система костных устроена несколько сложнее, так как им постоянно приходится поддерживать разницу в солевом составе тела и среды.
Почки пресноводных костных рыб вынуждены постоянно реабсорбировать ионы (возвращать в организм из первичной мочи), иначе тело рыбы потеряет все соли. Конечный продукт азотистого обмена у пресноводных – не мочевина, а токсичный аммиак. Морские виды, наоборот, стремятся вывести соли и не потерять воду. Они выделяют гораздо меньше мочи, чем пресноводные.
У некоторых видов имеется мочевой пузырь, где скапливается моча. Жабры и кожа также могут участвовать в выведении продуктов обмена.
Нервная система
Головной мозг устроен проще, чем у хрящевых. Строение органов чувств схожее.
Поведение костных рыб сложнее. Если у акул рефлексы вырабатываются медленно и затухают за 2-3 дня, у костных рыб рефлекс может затухать в течении месяцев.
Половая система
Не сопряжена с выделительной, семявыносящие протоки самостоятельно открываются половым отверстием.
Оплодотворение у большинства видов наружное. Самка мечет яйца (икру), самец оплодотворяет её. Выбор партнёра и определение места для нереста у некоторых рыб может быть сложным процессом. Часто родители заботятся о потомстве: строят надёжные укрытия, присматривают за мальками.
Одна самка может метать тысячи икринок (рыба-луна – до 30 млн.). Икринки содержат много желтка, поэтому другие животные часто лакомятся ими.
дыхательная система | Определение, органы, функции и факты
Дыхательная система , система живых организмов, которая поглощает кислород и выделяет углекислый газ для удовлетворения энергетических потребностей. В живом организме энергия высвобождается вместе с углекислым газом в результате окисления молекул, содержащих углерод. Термин дыхание обозначает обмен дыхательных газов (кислорода и углекислого газа) между организмом и средой, в которой он живет, а также между клетками тела и тканевой жидкостью, которая их омывает.
дыхание: животные
Различные методы дыхания у животных.
Encyclopædia Britannica, Inc.
За исключением энергии, используемой животными в глубинах океана, вся энергия, используемая животными, в конечном итоге происходит из энергии солнечного света. Углекислый газ в атмосфере в сочетании с энергией солнечного света используется растениями для синтеза сахаров и других компонентов. Животные потребляют растения или другой органический материал для получения химических соединений, которые затем окисляются для поддержания жизненно важных процессов.
В данной статье рассматриваются газообразные компоненты воздуха и воды, естественные дыхательные среды обитания животных и основные типы дыхательных структур, которые способствуют газообмену в этих средах.
Хотя получение кислорода и удаление углекислого газа являются важными требованиями для всех животных, скорость и количество газообмена варьируются в зависимости от вида животного и степени его активности. В таблице потребление кислорода различными животными выражено в миллилитрах кислорода на килограмм веса тела в час, что отражает потребность в газе различных видов животных в состоянии покоя и в движении.Изменение химического состава жидкостей организма вызывает реакцию центральной нервной системы, которая затем возбуждает или подавляет механизмы внешнего дыхания.
Получите эксклюзивный доступ к контенту из нашего первого издания 1768 с вашей подпиской.
Подпишитесь сегодня
| животное | вес (грамм) | Потребление кислорода (миллилитры на килограмм веса в час) |
|---|---|---|
| Источник: А.Крог, Сравнительная физиология респираторных механизмов (1959). | ||
| парамеций | 0,000001 | 500 |
| мидия (Mytilus) | 25 | 22 |
| раки (Astacus) | 32 | 47 |
| бабочка (Ванесса), отдыхает | 0,3 | 600 |
| бабочка (Ванесса) летающая | 0,3 | 100 000 |
| карп (Cyprinus) | 200 | 100 |
| щука (Esox) | 200 | 350 |
| мышь в состоянии покоя | 20 | 2,500 |
| мышь, работает | 20 | 20 000 |
| человек в состоянии покоя | 70 000 | 200 |
| человек, максимальная работа | 70 000 | 4 000 |
Газы в окружающей среде
Диапазон респираторных заболеваний, с которыми сталкиваются водные и наземные животные, можно увидеть по разному составу и физическим характеристикам воды и воздуха.В воздухе содержится примерно в 20 раз больше кислорода, чем в насыщенной воздухом воде. Чтобы извлечь эквивалентное количество кислорода в качестве дышащего воздуха, водное животное может счесть необходимым пропускать через респираторные поверхности относительно больший объем внешней среды. Более того, скорость диффузии кислорода в воде намного ниже, чем в воздухе. Проблема еще больше усугубляется более высокой плотностью (в 1000 раз больше воздуха) и вязкостью (в 100 раз больше воздуха) воды, которые создают гораздо большую рабочую нагрузку на механизмы водного дыхания.Таким образом, рыба может расходовать около 20 процентов своего общего потребления кислорода на работу дыхательного насоса по сравнению с 1-2 процентами у млекопитающих, включая человека.
Содержание углекислого газа в большинстве природных вод низкое по сравнению с воздухом, часто почти нулевое. В отличие от кислорода, углекислый газ очень хорошо растворяется в воде и быстро диффундирует. Большая часть углекислого газа, попадающего в воду, соединяется либо с водой (с образованием угольной кислоты), либо с другими веществами (с образованием карбонатов или бикарбонатов).Эта буферная способность поддерживает низкий уровень свободного диоксида углерода и способствует поддержанию благоприятного градиента диффузии для обмена диоксида углерода водными сапунами. В общем, кислородный обмен, который сильно зависит от содержания кислорода в воде, является более критически ограничивающим для водных форм, чем обмен диоксида углерода.
Температура оказывает сильное влияние на растворимость газов в воде. Изменение с 5 ° до 35 ° C (от 41 ° до 95 ° F) снижает содержание кислорода в пресной воде почти вдвое.В то же время повышение температуры тела приводит к увеличению потребления кислорода животными, которые плохо регулируют температуру своего тела (так называемые хладнокровные животные). Рыба, испытывающая одновременно повышение температуры воды и тела, испытывает двойной недостаток: через ее жаберные поверхности необходимо прокачать больше воды, чтобы извлечь такое же количество кислорода, какое требуется при более низкой температуре; а усиленный метаболизм требует большего количества кислорода.
Количество кислорода, доступного в природных водах, также ограничено количеством растворенных солей.Этот фактор является определяющим фактором доступности кислорода в переходных зонах между морской и пресной водой. Чистая вода, когда она уравновешена кислородом при 0 ° C, например, содержит около 50 миллилитров кислорода на литр; при тех же условиях раствор, содержащий 2,9 процента хлорида натрия, содержит всего 40 миллилитров кислорода на литр. В водоемах могут быть зоны с низким содержанием кислорода. Такие зоны особенно заметны на болотах и на нижних уровнях глубоких озер. Многие животные исключены из таких зон; другие стали замечательно приспособлены к жизни в них.
Атмосфера Земли простирается на многие мили. Он состоит из смеси газов, удерживаемых в оболочке вокруг земного шара за счет гравитационного притяжения. Атмосфера оказывает давление, пропорциональное весу столба воздуха над поверхностью Земли, простирающегося до предела атмосферы: атмосферного давления на уровне моря в среднем достаточно, чтобы поддерживать столб ртути высотой 760 миллиметров (сокращенно 760 мм рт. Ст. (Последнее является химическим символом ртути).Сухой воздух состоит в основном из азота и инертных газов (79,02 процента), кислорода (20,94 процента) и углекислого газа (0,03 процента), каждый из которых пропорционально влияет на общее давление. Эти проценты относительно постоянны на высоте около 80,5 км. На уровне моря и барометрическом давлении 760 миллиметров ртутного столба парциальное давление азота составляет 79,02 процента от 760 миллиметров ртутного столба, или 600,55 миллиметра ртутного столба; кислород – 159,16 миллиметра ртутного столба; а диоксида углерода – 0.20 миллиметров ртутного столба.
Наличие водяного пара в газовой смеси снижает парциальные давления других составляющих газов, но не изменяет общее давление смеси. Важность давления водяного пара для состава газа можно оценить по тому факту, что при температуре тела человека (37 ° C или 98,6 ° F) атмосферный воздух, втягиваемый в легкие, становится насыщенным водяным паром. Давление водяного пара при 37 ° C составляет 47 миллиметров ртутного столба. Чтобы рассчитать парциальное давление дыхательных газов, это значение необходимо вычесть из атмосферного давления.Для кислорода 760 (атмосферное давление) – 47 = 713 миллиметров ртутного столба, а 713 × 0,209 (процентное содержание кислорода в атмосфере) = 149 миллиметров ртутного столба; это примерно на 10 миллиметров ртутного столба ниже, чем парциальное давление кислорода в сухом воздухе при общем давлении 760 миллиметров ртутного столба.
Атмосферное давление падает на больших высотах, но состав атмосферы остается неизменным. На высоте 7600 метров (25000 футов) атмосферное давление составляет 282 миллиметра ртутного столба, а парциальное давление кислорода составляет около 59 миллиметров ртутного столба.Кислород по-прежнему составляет лишь 20,94 процента от общего количества газа. Разряженность воздуха на больших высотах не только ограничивает доступность кислорода для дыхательного аппарата, но также ограничивает его доступность для водных форм, поскольку количество растворенного в воде газа уменьшается параллельно с падением атмосферного давления. Озеро Титикака в Перу находится на высоте около 3810 метров; в одном литре озерной воды на этой высоте (и при 20 ° C или 68 ° F) содержится четыре миллилитра кислорода в растворе; на уровне моря вмещает 6.4.
Различия в характеристиках воздуха и воды указывают на множество проблем, с которыми дыхательная система животных должна справляться, добывая достаточно кислорода для поддержания жизни.
Сравнительная патогеномика бактерий, вызывающих инфекционные заболевания рыб
Рыбы, живущие в дикой природе, а также выращиваемые в аквакультурных хозяйствах, восприимчивы к инфекционным заболеваниям, вызываемым филогенетически разнообразным набором бактериальных патогенов. Варианты контроля и лечения с использованием вакцин и лекарств неадекватны, неэффективны или практически неосуществимы.Классический подход к изучению бактериальных патогенов рыб рассматривает отдельные или несколько факторов вирулентности. В последнее время секвенирование генома ряда бактериальных патогенов рыб значительно расширило наше понимание биологии, адаптации хозяина и факторов вирулентности этих важных патогенов. В этой статье делается попытка собрать разрозненную литературу по информации о последовательности генома патогенных бактерий рыб, опубликованную и доступную на сегодняшний день. Секвенирование генома выявило несколько сложных адаптивных эволюционных стратегий, опосредованных горизонтальным переносом генов, вставками элементов последовательности, мутациями и последовательностями профагов, действующих в патогенах рыб, и тем, как их геномы эволюционировали из универсальных экологических штаммов в высоковирулентные обязательные патогены.Кроме того, сравнительная геномика позволила идентифицировать уникальные кластеры генов, специфичных для патогенов. Статья посвящена сравнительному анализу вирулогеномов важных бактериальных патогенов рыб и генов, участвующих в их эволюционной адаптации к различным экологическим нишам. В статье также предлагаются некоторые новые направления поиска новых вакцин и химиотерапевтических мишеней в геномах бактериальных патогенов рыб.
1. Введение
Секвенирование генома дало нам глубокое понимание генетической структуры микробного мира.Сегодня микробная геномика перешла от давно затянувшихся проектов секвенирования отдельных геномов в прошлом до уровня технического прогресса, когда секвенирование и сравнение геномов нескольких штаммов одного патогена выполняется за очень короткий период времени [1, 2 ]. В настоящее время мы переживаем период бурного развития в этой области и подавляющего избытка данных о последовательности генома микроорганизмов. На сегодняшний день опубликовано более 1800 микробных геномов, и секвенирование более 5200 микробных геномов находится на разных стадиях завершения (http: // www.ncbi.nlm.nih.gov/genomes/lproks.cgi).
Информация о геномике категорически опровергла прежнее представление о том, что микробные геномы статичны, и продемонстрировала, что процессы геномной эволюции намного более гибкие и динамичные, чем считалось ранее. Это привело к появлению новых идей, таких как «искоренение древа жизни» и концепции «горизонтальной геномики» [3–8]. Это новое представление об эволюции микробного генома возникло из наблюдений за редукцией генома, специфичной для клонов, и горизонтальным переносом генов (ГПГ), часто встречающимся в бактериальных геномах.Все чаще проекты по секвенированию генома выявляют неожиданный уровень разнообразия среди бактерий, которое часто может быть связано с рекомбинацией и переносом генов между различными прокариотическими организмами.
Размеры и состав бактериальных геномов сильно различаются между разными родами и видами, а также между штаммами одного и того же вида. Известные размеры генома бактерий варьируются от 0,6 до 10 мегабаз (МБ). Наименьшие бактериальные геномы зарегистрированы для микоплазм и родственных бактерий, их размер составляет всего 530 килобаз [9].Подчеркивается, что адаптивная способность («универсальность») бактерий напрямую коррелирует с размером генома [10].
Секвенирование генома бактериальных патогенов дало захватывающую информацию об эволюционных взаимоотношениях между патогенными и непатогенными видами и продемонстрировало, как каждый из них развил особые адаптации, выгодные для каждого из их уникального инфекционного образа жизни. В более долгосрочной перспективе понимание их генома и биологии позволит ученым разработать средства нарушения их заразного образа жизни.
Геномы бактерий состоят из круговых или линейных хромосом, внехромосомных линейных или кольцевых плазмид, а также из различных комбинаций этих молекул. Функционально связанные гены сгруппированы вместе в очень непосредственной близости друг от друга, и эти гены, расположенные в «основной» части хромосомы, представляют относительно однородное содержание G + C и конкретное использование кодонов. Близкородственные бактерии обычно имеют очень похожие геномы [11].
Однако стабильность и целостность «основных» последовательностей генома часто нарушается из-за присутствия фрагментов ДНК с содержанием G + C и использованием кодонов, заметно отличающимся от таковых в «стержневом» геноме.«Гибкий» генофонд, или так называемый «мобилом» [12], создается путем получения специфических для штаммов «ассортимента» генетической информации, в основном представленной мобильными генетическими элементами (MGE), такими как плазмиды, бактериофаги, геномные / острова патогенности (GEIs / PAIs), интегроны, IS-элементы (ISEs) и транспозоны (см. рисунок 1). Гибкие гены, разбросанные по геному, обеспечивают микробам дополнительный арсенал, например устойчивость к антибиотикам, выработку токсичных соединений, а также других факторов вирулентности [13].
Фундаментальный вопрос биологии – определить минимальное количество генов или функций, поддерживающих клеточную жизнь. Размер бактериальных геномов в первую очередь является результатом двух противодействующих процессов: приобретения новых генов путем дупликации генов или путем горизонтального переноса генов; удаление несущественных генов. Геномный поток, создаваемый этими приобретениями и потерями генетической информации, может существенно изменить содержание генов. Этот процесс приводит к дивергенции видов бактерий и, в конечном итоге, к адаптации к новым экологическим нишам [16].
Бактериальные патогены являются основной причиной инфекционных заболеваний и смертности диких рыбных запасов и рыб, выращиваемых в замкнутых условиях. Проблемы, связанные с болезнями, являются самой крупной причиной экономических потерь в аквакультуре [17]. Одновременно с быстрым ростом и интенсификацией аквакультуры, увеличением использования водоемов, загрязнением, глобализацией и трансграничным перемещением водной фауны постоянно увеличивается список новых патогенных видов бактерий, выделенных из рыб [18].Кроме того, вирулентность и круг хозяев существующих патогенов также увеличиваются, что создает серьезную проблему для исследователей здоровья рыб, которые активно ищут более эффективные вакцины и терапевтические препараты для борьбы с бактериальными болезнями рыб. Современные методы лечения малоэффективны и имеют много практических трудностей.
На уровне взаимодействия между хозяином и патогеном патогены испытывают значительное давление, чтобы они адаптировались к суровой среде хозяина, а также адаптировались и развивались вместе с постоянно меняющейся внешней средой.Взаимодействие между хозяином и патогеном является сложным, и каждое из них обусловлено необходимостью обеспечить успех вида. Адаптация одного партнера к использованию новой среды часто стимулирует другого к изменению ее характеристик, чтобы воспользоваться этим изменением. В результате этого цикла взаимодействия, созданного изменяющейся окружающей средой, будут развиваться новые штаммы патогенов. Со временем эти штаммы могут появиться как новые виды с характерными симптомами болезни.
Использование антибиотиков для борьбы с болезнями рыб имеет ограниченный успех и представляет потенциальную опасность развития устойчивости к антибиотикам у водных бактерий (фактологический бюллетень Всемирной организации здравоохранения по устойчивости к противомикробным препаратам 194, http: // www.who.int/inf-fs/en/fact194.html) [19]. Поскольку аквакультура является одной из самых быстрорастущих отраслей производства продуктов питания в мире, потребность в устойчивых способах борьбы с болезнями рыб становится все более актуальной. Существуют огромные возможности для разработки новых вакцин и терапевтических препаратов против бактериальных патогенов рыб.
Геномная эволюция и стратегии адаптации бактериальных патогенов рыб плохо изучены и сильно отстают от патогенов человека и наземных животных. Детальное знание последовательностей генома бактериальных патогенов рыб и того, как геномы патогенных видов или штаммов произошли от непатогенных предков или аналогов, поможет нам лучше понять механизмы их патогенности и стратегии адаптации хозяев.Эта информация поможет идентифицировать новые вакцины и лекарства-мишени в геномах патогенов.
Недавно было завершено секвенирование генома ряда бактерий, патогенных для рыб и других водных организмов. Последовательность генома и характеристики генома важных бактериальных патогенов рыб, составленные и опубликованные на сегодняшний день, сведены в Таблицу 1.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
* Не опубликовано. ** Кодирующие последовательности. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Основная цель данной статьи – собрать и обобщить разрозненную информацию о секвенировании генома важных бактериальных патогенов рыб, имеющуюся на сегодняшний день в литературе. Мы искренне верим, что эта статья предоставит геномный взгляд на адаптивные эволюционные стратегии бактериальных патогенов рыб в различных экологических нишах и поможет лучше понять механизмы вирулентности и патогенез инфекций.Есть надежда, что это приведет к поиску наиболее подходящих мишеней для вакцин и терапевтических препаратов в геномах и разработке эффективных методов контроля и лечения болезней рыб.
2. Бактериальные патогены рыб
Хотя патогенные виды, представляющие большинство существующих бактериальных таксонов, причастны к болезням рыб, только относительно небольшое количество патогенов несет ответственность за значительные экономические потери выращиваемой рыбы во всем мире. Основные бактериальные патогены, ответственные за вспышки инфекционных заболеваний у разных видов рыб, перечислены в таблице
Роль Nrf2 и его активаторов в респираторных заболеваниях
Фактор транскрипции ядерный фактор Эритроид 2 Фактор 2 (Nrf2) является основным регулятором антиоксидантной реакции элемент- (ARE-) управляемая экспрессия цитопротекторного белка.Активация передачи сигналов Nrf2 играет важную роль в предотвращении повреждений клеток и тканей, вызванных окислительным стрессом. В условиях отсутствия стресса природный ингибитор Nrf2, Kelch-подобный ECH-ассоциированный белок 1 (Keap1), улавливает Nrf2 в цитоплазме и способствует деградации Nrf2 протеасомой 26S. Тем не менее, стрессы, включая высокоокислительные микроокружения, нарушают способность Keap1 нацеливаться на Nrf2 для убиквитинирования и деградации и побуждают недавно синтезированный Nrf2 перемещаться в ядро для связывания с ARE.Из-за постоянного воздействия внешней среды, включая различные загрязняющие вещества и другие окислители, окислительно-восстановительный баланс, поддерживаемый Nrf2, довольно важен для дыхательных путей. На сегодняшний день исследователи обнаружили, что делеция Nrf2 приводит к высокой восприимчивости и серьезности инсультов в различных моделях респираторных заболеваний, включая бронхолегочную дисплазию (БЛД), респираторные инфекции, острый респираторный дистресс-синдром (ОРДС), хроническую обструктивную болезнь легких (ХОБЛ), астма, идиопатический фиброз легких (IPF) и рак легких.Напротив, активация Nrf2 оказывает защитное действие на эти заболевания легких. В настоящем обзоре мы суммируем участие Nrf2 в патогенезе вышеуказанных респираторных заболеваний, которые были идентифицированы экспериментальными моделями и исследованиями на людях, и описываем защитные эффекты индукторов Nrf2 в отношении этих заболеваний.
1. Окислительный стресс и антиоксидантная реакция при респираторных заболеваниях
За последние несколько десятилетий экологические проблемы, вызванные антропогенными и природными факторами, резко увеличили заболеваемость злокачественными и незлокачественными респираторными заболеваниями.Поэтому причина, лежащая в основе того, почему на дыхательную систему так легко влияют экологические проблемы и патогенез респираторных заболеваний, привлекает все большее внимание.
Являясь местом газообмена, дыхательные пути с большой площадью поверхности постоянно контактируют с внешней средой и подвергаются воздействию различных токсичных веществ, переносимых по воздуху, особенно вдыхаемых окислителей (например, озона окружающей среды, частиц и сигаретного дыма) [1]. В связи с особенностями анатомии и физиологии дыхательные пути помещены в высокоокислительную микросреду.Следовательно, окислительно-восстановительный гомеостаз в дыхательных путях может быть легко нарушен, что называется окислительным стрессом [2]. Окислительный стресс – это обычное состояние, определяемое как дисбаланс между производством активных форм кислорода (АФК) и антиоксидантной способностью клеток при временной или постоянной стимуляции обильными оксидантными стрессорами [3]. Недавно было доказано, что оксидативный стресс связан с патогенезом различных острых и хронических респираторных заболеваний, включая респираторные инфекции, острый респираторный дистресс-синдром (ОРДС), хроническую обструктивную болезнь легких (ХОБЛ), астму, идиопатический фиброз легких (IPF), и рак легких [4–6].В легких людей с этими заболеваниями всегда наблюдается нарушение окислительно-восстановительного баланса, которое может быть представлено повышенными биомаркерами окислительного стресса.
Тем не менее, во время долгого пути эволюции жизни организм развил серию антиоксидантных реакций, противодействующих токсичности окислительного стресса. Антиоксидантная система в клеточном ответе включает либо белки (например, ферменты), либо небольшие молекулы (например, витамины C и E). Поскольку было доказано, что ферменты играют важную роль в жизненных циклах, их влияние на антиоксидантную защиту было тщательно исследовано.Прямые антиоксидантные ферменты относятся к классическим ферментам, включая супероксиддисмутазы (SOD), каталазу и глутатионпероксидазу (GPx), тогда как непрямые антиоксидантные ферменты в основном относятся к детоксифицирующим ферментам фазы 2, таким как изоферменты глутатион-S-трансферазы (GST), каталитические и модифицирующие субъединицы. γ -глутамилцистеинлигазы (GCLC, GCLM) и НАДФ (H): хиноноксидоредуктаза (NQO1). Более того, сообщается, что гемоксигеназа (HO-1), отвечающая за стрессовую реакцию, является особенно мощным антиоксидантным белком [7–9].Антиоксидантные вещества присутствуют в относительном количестве как в эпителиальной подкладочной жидкости (ELF), так и в тканях легких, поскольку дыхательные пути являются местами, где обычно происходят реакции детоксикации, и защищают легкие от окислительных повреждений у здоровых людей [2]. К сожалению, у восприимчивых людей может происходить истощение GSH и других антиоксидантов, и эти люди склонны к развитию окислительных респираторных заболеваний. Более того, защитная роль антиоксидантной системы в предотвращении этих респираторных заболеваний также может быть доказана несколькими методами лечения, направленными на защиту от окислительного стресса, которые уже применялись при БЛД, ХОБЛ и ИПФ, например, лечение витаминами С и Е. или N-ацетилцистеин (NAC, предшественник GSH) или обработка конъюгированными с полиэтиленгликолем SOD и каталазой.
2. Nrf2-опосредованный антиоксидантный путь и респираторные заболевания
Хотя функциональные механизмы в антиоксидантной системе различны, большое количество типичных детоксифицирующих ферментов фазы 2 и белка реакции на стресс HO-1 регулируются ядерным фактором транскрипционного фактора. erythroid 2-related factor 2 (Nrf2), что указывает на то, что этот фактор транскрипции является возможным и обязательным вышестоящим регулятором антиоксидантных ответов, который поддерживает клеточный окислительно-восстановительный гомеостаз и снижает тяжелое окислительное повреждение [10–13].
Nrf2, который принадлежит к семейству факторов транскрипции cap «n» воротник (CNC), является основным фактором транскрипции, который противодействует окислительному стрессу и воспалению посредством скоординированной индукции транскрипции цитопротекторного гена, управляемой элементом антиоксидантного ответа (ARE-) [ 14, 15]. Классические механизмы активации Nrf2 включают окислительную модификацию и конформационные изменения в его основном репрессорном белке, Kelch-подобном ECH ассоциированном белке 1 (Keap1), с последующей стабилизацией Nrf2 из-за выхода из убиквитинирования с помощью Cul3-Rbx1.Эта молекулярная модель подтверждается конститутивным накоплением Nrf2 в ядрах мышей с нокаутом по Keap1 [16]. Фактически, Nrf2 состоит из шести функциональных доменов, распознаваемых как гомологии Nrf2-ECH 1-6 (Neh2-6), включая связывающий домен Keap1 (Neh3) и домен лейциновой молнии (Neh2), через которые Nrf2 может гетеродимеризоваться с небольшими Maf или Белки Jun, а затем связываются с ARE [17]. Репрессор keap1 Nrf2 представляет собой цитоплазматический и богатый цистеином белок, N-концевой домен BTB которого связывается с Cullin 3- (Cul3-) Rbx1, тогда как C-концевой домен DGR связывается с Nrf2.Keap1 репрессирует Nrf2, выступая в качестве адаптера субстрата для Cul3-содержащего комплекса убиквитин-лигазы E3. В физиологических условиях Keap1 удерживает Nrf2 в цитоплазме и убиквитинирует Nrf2 для облегчения его деградации с помощью протеасомы 26S. Однако, когда существуют окислительные стимулы, остатки цистеина Keap1 могут быть модифицированы, что приводит к стабилизации и накоплению Nrf2 в ядре [18]. Несколько механизмов активации Nrf2 с помощью Keap1 были предложены до сих пор [17, 19, 20].(1) Диссоциация Keap1: модификация цистеина в Keap1 заставляет Nrf2 диссоциировать от Keap1. (2) Петля и защелка Keap1: как более широко принятая модель, Nrf2 связывается с гомодимером Keap1 посредством высокоаффинного мотива ETGE в качестве «шарнира» и низкоаффинного мотива DLG в качестве «защелки». Модификация цистеина в Keap1 приводит к конформационным изменениям, но не запускает диссоциацию Nrf2, которая может ингибировать связывание убиквитина с Nrf2, нарушая сайт связывания слабой защелки.(3) Убиквитинирование Keap1: модификация цистеина в Keap1 перемещает конъюгацию убиквитина от Nrf2 к самому себе. Тем не менее, точные молекулярные механизмы, лежащие в основе того, как Nrf2 обходит ворота Keap1 в стрессовых условиях, все еще остаются невыясненными. Несколько исследований, проведенных в последние годы, предложили довольно интригующие теории. Напр., Cys-151 в домене BTB Keap1, вероятно, играет важную роль в ответ на активаторы Nrf2, и это действие может быть связано с его деструктивным действием на взаимодействие Keap1 с Cul3 [21].Разнообразные активаторы Nrf2 активируют передачу сигналов Nrf2 посредством этого канонического механизма, а мутация цистеина 151 в серин (Keap1-C151S) в Keap1 полностью устраняет повышающую регуляцию Nrf2 [22, 23]. Другие исследования показывают, что Cys-273 и Cys-288 в Keap1 могут также вносить вклад в структурную целостность и активность Keap1 для поддержания активности ubiquitin ligase [24]. Кроме того, существуют также независимые от Keap1 пути активации Nrf2, среди которых протеинкиназы играют важную роль.Предыдущее исследование показало, что фосфорилирование по определенному аминокислотному остатку Nrf2 может повысить его стабильность и трансактивационную активность [8]. Типичные пути протеинкиназ включают фосфатидилинозитол-3-киназу (PI3K), MAPK, PKC и киназу-3 гликогенсинтазы (GSK-3). Фосфорилирование Nrf2 с помощью PI3K, PKC, c-Jun, N-концевой киназы (JNK) и протеинкиназы, регулируемой внеклеточными сигналами (ERK), обеспечивает положительную регуляцию, тогда как p38 MAPK регулирует путь Nrf2 как положительно, так и отрицательно [19, 25– 29].В последнее время неканонический путь активации Nrf2, связанный с аутофагией, привлекает все большее внимание из-за его двойного эффекта. Этот путь тесно связан с секвестосомой 1 белка субстрата аутофагии (SQSTM1 или p62). p62 может конкурировать с Nrf2 за связывание Keap1, секвестрировать Keap1 в аутофагосому и обеспечивать стабилизацию и накопление Nrf2 [30]. Однако активация Nrf2, опосредованная аутофагией, имеет как положительные, так и отрицательные эффекты: индукция аутофагии приводит к секвестрации комплексов Keap1-p62 в аутофагосомы и опосредованной лизосомой деградации Keap1, что приводит к контролируемой активации Nrf2 и оказывает защитные эффекты, в то время как приводит к нарушению регуляции аутофагии. при длительной активации Nrf2 в патологическом состоянии и оказывает пагубное воздействие.
Более того, недавние исследования показали, что помимо Keap1 др. Пути с участием адаптерных белков cullin также направляют убиквитинирование Nrf2. Например, фосфорилирование Nrf2 по конкретным сериновым остаткам в домене Neh6 с помощью GSK-3 образует часть деградации для распознавания адаптером убиквитинлигазы Skp1-Cul1-F-box белок (SCF) / β -TrCP ( β -трансдуциновый повтор, содержащий белок) и деградацию протеасом комплексом Cullin1 / Rbx1 [31, 32].Точно так же WDR23, рецептор субстрата для Cullin4- (CUL4-) DDB1 (поврежденный ДНК-связывающий белок 1) E3-ubiquitin ligase, связывается с доменом Neh3 Nrf2 и негативно регулирует уровень и активность Nrf2 [33].
Функциональный процесс происходит, когда de novo синтезированный Nrf2 перемещается в ядро, после чего Nrf2 гетеродимеризуется с небольшими белками Maf или Jun, а затем связывается с ARE в регуляторных областях генов-мишеней Nrf2 и либо активирует, либо ингибирует гены-мишени.Предыдущие исследования показали, что в защите дыхательной системы с помощью Nrf2 гены, нацеленные на Nrf2, являются важными эффекторами, которые идентифицируются с помощью анализа микроматриц и биоинформатических исследований. Путь Nrf2 / ARE регулирует более 500 генов, включая гены, регулирующие окислительный стресс (HO-1, GCLM и GCLC), воспаление (TGF- β и NF- κ B), метаболизм и выделение ксенобиотиков (NQO1 , AKR1C1 и MRP1), апоптоз (Bcl-2 и BclxL) и аутофагия (p62) [12, 23].Следовательно, гены-мишени, расположенные ниже по течению Nrf2, выполняют разнообразные функции, включая антиоксидантную, противовоспалительную, метаболизм ксенобиотиков, детоксикацию и регуляцию роста клеток [25, 34, 35]. В частности, цитопротективные белки, такие как HO-1, GCLM, GCLC и NQO1, катализируют различные реакции детоксикации, превращая вредные вещества в гидрофильные метаболиты, и не потребляются во время их действия [36] (Рисунок 1). Более того, защитные эффекты Nrf2 с большей вероятностью проявляются, когда его активация строго контролируется.Недавнее исследование показало, что как только окислительно-восстановительный гомеостаз восстанавливается или соединения метаболизируются и элиминируются, происходит прекращение сигнала, и Keap1 входит в ядро, связывается с Nrf2, а затем возвращает его в цитозоль для деградации [37].
Соответственно, предыдущие исследования показали, что в тканях, где происходят обычные процессы антиоксидирования и детоксикации, таких как легкие, печень и почки, Nrf2 относительно широко распространен. Уровень экспрессии Nrf2 сильно коррелирует с восприимчивостью, серьезностью и восстановлением нарушений дыхательных путей.Данные, полученные на мышах с нокаутом Nrf2, идентифицируют защитные эффекты Nrf2 при нарушениях дыхательных путей: по сравнению с мышами дикого типа, мыши с нокаутом Nrf2 имеют усиленное воспаление легких, повреждение эпителиальных клеток и повышенную чувствительность к сигаретному дыму, эластазе, овальбумину, блеомицину и другие раздражители [38–41]. Эти результаты побуждают исследователей открывать потенциальные активаторы Nrf2.
3. Защитная роль Nrf2 и его активаторов при респираторных заболеваниях
Как описано выше, дыхательная система напрямую подвергается воздействию вдыхаемых оксидантов, и это окислительное бремя делает эту систему более уязвимой к окислительному стрессу, который, как было доказано, связан с патогенез разнообразных респираторных заболеваний.В настоящее время защитная роль передачи сигналов Nrf2 при респираторных расстройствах была идентифицирована с применением мышей с нокаутом Nrf2, включая три разных генетических фона (ICR, C57BL / 6J и Balbc / J), и мышей с условным нокаутом в легких [1]. Например, делеция Nrf2 связана с более серьезными поражениями, которые вызваны окислительными, воспалительными или канцерогенезными факторами, такими как инфекция, гипероксия, сигарета, овальбумин, блеомицин, бутилированный гидрокситолуол и частицы выхлопных газов дизельного топлива.Напротив, активация Nrf2 оказывает защитное действие на эти заболевания легких. Поэтому многие исследования сосредоточены на преимуществах индукторов Nrf2, включая натуральные продукты, в терапевтических вмешательствах при заболеваниях легких, связанных с оксидантами [42]. В настоящем обзоре мы обсуждаем участие Nrf2 в патогенезе заболеваний дыхательных путей и защитную роль различных индукторов Nrf2 при различных заболеваниях дыхательных путей. Кроме того, эффекты основных активаторов Nrf2 на различные респираторные заболевания сведены в таблицу (Таблица 1).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.1. Бронхолегочная дисплазия (БЛД)
Бронхолегочная дисплазия (БЛД) – это хроническое респираторное заболевание, которое обычно возникает у недоношенных детей с очень низкой массой тела при рождении. БЛД в основном характеризуется нарушением альвеоляризации, что приводит к нарушению роста альвеол и развития сосудов, воспалению легких и нарушению функции легких [43, 44]. Более того, ПРЛ не только приводит к значительной смертности в перинатальном периоде, но также способствует долгосрочным последствиям у подростков или раннего взросления с клинически значимыми респираторными симптомами [45].
Хотя патогенез ПРЛ до конца не изучен, повреждение легких при ПРЛ можно разделить на две группы, описанные как «новые» и «старые» ПРЛ. Первое относится к ранней остановке развития из-за пренатального воздействия или генетических факторов, тогда как второе приводит к появлению недоношенных детей с дефицитом сурфактанта, которым оказывается респираторная поддержка. Дальнейшие исследования выясняют, что важные триггерные события для ПРЛ могут включать окислительное повреждение и воспаление. В частности, когда человеческие младенцы претерпевают критический постнатальный альвеолярный рост, терапевтически вводимый кислород для улучшения оксигенации недоношенных детей является основным фактором риска, который может быть связан с индукцией регуляторных генов p21 и p53 [46].При окислительных расстройствах у взрослых было доказано, что активация Nrf2 полезна; тем не менее, исследование его воздействия на неонатальные заболевания все еще остается новой областью. Как свидетельствуют недавние исследования, в процессе созревания мешковидных легких Nrf2 может модулировать различные гены, участвующие в поддержании окислительно-восстановительного баланса, развитии органов и морфогенезе легких, а также росте и гибели клеток, а также иммунитете. Действительно, предыдущие исследования указывают на влияние Nrf2 на молекулярные процессы альвеоляризации и заболевания легких при преждевременных родах: дефицит Nrf2 и воздействие гипероксии у новорожденных мышей увеличивает смертность и серьезность ингибирования роста альвеол, а анализ транскриптома незрелой легочной ткани предполагает защита от O 2 токсичности Nrf2 может быть опосредована его регуляцией клеточного цикла, процессов метаболизма, межклеточных взаимодействий и окислительно-восстановительного гомеостаза [47, 48].
Эти данные могут пролить свет на возможную полезную роль активаторов Nrf2 в ПРЛ у недоношенных новорожденных в отношении как развития легких, так и гипероксии-опосредованного повреждения легких, и несколько исследований попытались раскрыть такой потенциал. Например, ауротиоглюкоза (ATG), ингибитор TrxR1, ингибирует активность тиоредоксинредуктазы-1 (TrxR1) и активирует путь Nrf2 в легких новорожденных мышей C3H / HeN, подвергшихся гипероксии, ослабляя снижение массы тела и изменения в развитии альвеол. .Однако эффекты на альвеоляризацию и устойчивую активацию Nrf2 не наблюдаются в легких новорожденных мышей C57BL / 6 в тех же условиях [49, 50]. В модели БЛД у новорожденных крыс активатор Nrf2 куркумин может ослаблять гипероксическое повреждение легких, и считается, что защитная роль по крайней мере частично связана с активацией Nrf2; К сожалению, не было изучено, активируется ли путь Nrf2 [51, 52]. Другое исследование показывает, что внутриутробное введение хорошо известного индуктора Nrf2, сульфорафана, может ингибировать вызванное гипероксией воспаление легких у новорожденных мышей, но подавление альвеолярного роста не улучшается в этой модели БЛД [53].Такие результаты являются провокационными, и вопрос о том, является ли индукция Nrf2 полезной в текущих условиях при БЛД, может быть спорным, и необходимы исследования долгосрочного воздействия индукторов Nrf2 in utero на альвеолярный рост, воспаление и выживаемость. Принимая во внимание эти открытия, активация Nrf2 может быть новой стратегией в предотвращении или модуляции тяжести ПРЛ, а также в улучшении исходов болезни. Однако следует проявлять осторожность при оценке эффектов индукторов Nrf2 при БЛД перед клиническим применением, поскольку результаты предыдущих исследований все еще ограничены и противоречивы в некоторых отношениях, таких как способ введения, время доставки и даже вид.
3.2. Респираторные инфекции
Поскольку дыхательные пути напрямую контактируют с различными патогенами из внутренней и внешней среды, связанные с ними инфекции широко распространены и разнообразны. Таким образом, все еще существует неудовлетворенная потребность в новых терапевтических методах лечения этих заболеваний, особенно вирусных инфекций. Недавние исследования показывают, что во время процесса респираторных инфекций чрезмерное количество АФК, продуцируемых фагоцитарными клетками, обычно вызывает дисбаланс между оксидантами и антиоксидантами, что может способствовать патогенезу респираторных заболеваний, связанных с инфекцией.Считается, что Nrf2 является важным борцом с окислительным стрессом, он также играет защитную роль в противовирусных и антибактериальных процессах при некоторых распространенных инфекциях дыхательных путей.
3.2.1. Инфекция, вызванная респираторно-синцитиальным вирусом (RSV)
Респираторно-синцитиальный вирус (RSV) является не только ведущей причиной острых респираторных инфекций у младенцев и детей, но также важным фактором значительной респираторной заболеваемости и смертности среди пожилых людей [54, 55].Однако детали, касающиеся патогенеза RSV-инфекции, не выяснены, а эффективные вакцины или лечение все еще недоступны. В недавних исследованиях исследователи обнаружили, что быстро генерируемые ROS во время RSV-инфекции участвуют в воспалительном и окислительном повреждении легких при этом клиническом заболевании [56]. Соответственно, лечение антиоксидантами может облегчить вызванное RSV воспаление легких на мышиной модели RSV-инфекции [57]. Изучая антиоксидантную активность передачи сигналов Nrf2, исследователи также обнаружили активацию Nrf2 во время инфекции RSV.Эти результаты показывают, что инфекция RSV вызывает явное снижение уровней Nrf2 и антиоксидантных ферментов в дыхательных путях в легких мышей, а также в выделениях носоглотки у детей, что может быть связано с индуцированной RSV деградацией Nrf2 [58–61]. Инфекция RSV способствует убиквитинированию и деградации Nrf2 через протеасомный путь, что может быть правдоподобным, поскольку протеасомные ингибиторы MG132 и лактацистин могут восстанавливать активность Nrf2, сниженную RSV. Что еще более важно, фенотип RSV-инфекции более серьезен у мышей с дефицитом Nrf2, чем у мышей дикого типа, что отражается в более тяжелом бронхолегочном воспалении и повреждении эпителия, а также в снижении клиренса вируса [62].Исследования показывают, что более тяжелое воспаление дыхательных путей у мышей Nrf2 – / – , вызванное RSV, вероятно, связано с усиленной одновременной активацией AP-1 и NF- κ B. Кроме того, дефицит Nrf2 делает мышей более «Th2-подобными». »С более низкими уровнями GSH и IFN- γ , что может ухудшить клиренс вируса. Следовательно, путь Nrf2 может быть направлен на защиту от повреждения легких, вызванного RSV.
В последние годы было обнаружено, что несколько индукторов Nrf2 оказывают положительное влияние на инфекцию RSV.Предварительная обработка сильнодействующим индуктором Nrf2 сульфорафаном значительно ограничивает репликацию RSV в легких и острое воспаление у мышей дикого типа, но не у мышей с дефицитом Nrf2 [62]. Бутилированный гидроксианизол (BHA), соединение, которое, как известно, индуцирует путь Nrf2, может ускорять клиренс вируса и уменьшать вызванное RSV воспаление легких у мышей, что указывает на то, что анти-RSV активность BHA может быть результатом активации Nrf2 [57 , 63]. Кроме того, ингибитор фосфодиэстеразы 4 (PDE4), рофлумиласт N-оксид (RNO), может обратить вспять вызванную RSV потерю Nrf2, и такие эффекты могут быть связаны с его ингибированием активности ресничек, продукции муцина и воспалительных медиаторов при RSV-инфекции в лунке. -дифференцированные нормальные эпителиальные клетки бронхов человека (WD-HBE) [64].Точно так же индуктор Nrf2 трет-бутилгидрохинон (tBHQ) также может восстанавливать снижение активации Nrf2, вызванное RSV [61].
3.2.2. Инфекция, вызванная вирусом гриппа A (IAV)
Инфекция, вызванная вирусом гриппа A (IAV), от инфекции верхних дыхательных путей до пневмонии, беспокоит людей в течение долгого времени [65, 66]. Несмотря на то, что были разработаны и применяются некоторые противовирусные препараты, такие как ингибиторы нейраминидазы (NA), инфекция IAV по-прежнему представляет собой серьезную угрозу для здоровья человека со значительной заболеваемостью и смертностью из-за вариабельности вируса и устойчивых вирусных штаммов, включая h2N1, H5N1 и H7N9 [67–69].Таким образом, разработка новых противогриппозных препаратов для борьбы с эпидемиями и пандемиями гриппа в будущем по-прежнему является важной задачей. В последние десятилетия исследования показали, что вирусы гриппа могут вызывать окислительный стресс, цитотоксичность, апоптоз и воспаление в дыхательной системе [70, 71]. Соответственно, антиоксидантные соединения защищают от повреждений, вызванных IAV, путем ингибирования репликации вируса и иммунного клиренса и уменьшения продукции медиатора воспаления [72]. Более того, антиоксидантный путь, контролируемый Nrf2, оказался центральным в антиоксидантной защите легких против воспаления и повреждения, вызванного вирусом гриппа, как в эпителиальных клетках носа человека, так и у мышей [73, 74].Исследования также показали, что во время инфекции IAV Nrf2 защищает клетки альвеолярного эпителия человека от цитопатических эффектов, вызванных окислительным стрессом, интерферон-независимым образом. Нокдаун Nrf2 делает ATI-подобные клетки и ATII клетки более уязвимыми к повреждению, в то время как избыточная экспрессия Nrf2 снижает репликацию вируса и связанную с ним экспрессию нуклеопротеинов [75]. Однако в будущих исследованиях потребуются клетки из других уязвимых групп населения, включая курильщиков и пациентов с ХОБЛ.
На сегодняшний день несколько индукторов Nrf2 были исследованы на предмет их защитной роли при инфекции IAV. Недавно сообщалось, что эпигаллокатехин галлат, индуктор Nrf2, снижает проникновение и репликацию вируса гриппа A / Bangkok / 1/79 в эпителиальных клетках носа; однако ингибирующий эффект на репликацию блокируется, когда Nrf2 сбивается [73], тогда как куркумин, классический индуктор Nrf2, не только инактивирует IAV и напрямую подавляет адсорбцию IAV, но также индуцирует активацию Nrf2 и ингибирует индуцированный IAV окислительный стресс, воспаление, и даже репликация IAV [76].Точно так же индукторы Nrf2 сульфорафан и эмодин также могут приводить к значительному ингибированию индуцированного IAV окислительного стресса / воспаления и репликации вируса [73, 77]. Другие сообщаемые соединения, которые могут оказывать противогриппозное действие посредством активации Nrf2, также включают бакучиол и производное рупестоновой кислоты, YZH-106 [67, 78].
3.2.3. Бактериальные инфекции
Недавно сообщалось, что Nrf2 играет полезную роль в инфицировании различными бактериями, и применение индукторов Nrf2 может представлять собой новое лечение бактериальных инфекций дыхательных путей, хотя связанные с этим исследования ограничены.
Стрептококк – важная бактерия, которая колонизирует верхние дыхательные пути. Эта бактерия является не только наиболее частой причиной внебольничной пневмонии, но и важным фактором риска ряда опасных для жизни заболеваний, включая сепсис [79]. Предыдущие исследования показали, что S. pneumoniae может высвобождать несколько бактериальных компонентов, вызывая повышенную окислительную нагрузку на эпителиальную поверхность легких, в то время как специфический индуктор Nrf2 ресвератрол может уменьшать вызванный пневмококком окислительный стресс в эпителии дыхательных путей [80].Аналогичным образом, Nrf2 может также эффективно ослаблять повреждение легких у мышей и уровень смертности, вызванный Staphylococcus aureus и воспалением легких при инфекциях Haemophilus influenzae [81, 82]. Более того, предварительная активация Nrf2 нуклеосомно-связывающим белком 2 группы с высокой подвижностью (HMGN2) может ослаблять окислительный стресс, вызванный инфекцией, вызванной Pseudomonas aeruginosa (PA), и ингибировать интернализацию бактерий в клетках A549 [83]. К сожалению, до сих пор нет данных о применении активаторов Nrf2 при этих инфекциях.
Более того, туберкулез (ТБ) является типичным примером конкретной инфекции и главным убийцей инфекционных заболеваний во всем мире. К туберкулезу в основном относятся инфекции, вызываемые Mycobacterium tuberculosis , особенно в дыхательных путях [84, 85]. В последнее время все большее внимание уделяется роли оксидантного стресса и Nrf2 в инфицировании туберкулезом. Было доказано, что оксидативный стресс существует на модели туберкулеза у морских свинок вместе с потерей активации Nrf2, а введение NAC, поглотителя ROS и активатора Nrf2, значительно снижает бактериальную нагрузку и повреждение легких при инфекции ТБ, указывая на возможное защитное действие. роль для Nrf2 [86].
3.3. Синдром острого респираторного дистресса (ОРДС)
Острый респираторный дистресс-синдром (ОРДС) относится к тяжелому клиническому состоянию одышки, рефрактерной гипоксемии и некардиогенного отека легких, которым страдают миллионы людей во всем мире. Согласно существующей статистике, этиология ОРДС довольно сложна, варьируется от тяжелой пневмонии, сепсиса и серьезной травмы до массивного переливания крови, причем тяжелая пневмония или сепсис могут быть основной причиной [87]. Тем не менее, типичные патологические изменения при ОРДС все еще имеют сходные характеристики, включая острое диффузное воспаление легких и нарушение эпителиально-сосудистого барьера, вызванное повреждением целостности эпителия и эндотелия [88].Эти изменения совместно приводят к разрушению структуры и функции легких, что в основном проявляется в нарушении эластичности легких и газообмена [89]. Несмотря на улучшение поддерживающей терапии сегодня, к сожалению, ОРДС по-прежнему является причиной значительной заболеваемости и смертности из-за неуловимых терапевтических методов.
Хотя механизмы, ответственные за патогенез ОРДС, до конца не изучены, уже есть данные, указывающие на участие двух основных и взаимосвязанных факторов, окислительного стресса и воспаления.Было доказано, что избыточное производство ROS или RNS, инфильтрация воспалительных клеток и синтез медиаторов воспаления вызывают серьезные повреждения легких. В последнее время появляется все больше данных, демонстрирующих важность активации Nrf2 в борьбе с окислительным стрессом и воспалением при ОРДС. Предыдущее исследование показало, что Nrf2 может выступать в качестве гена-кандидата восприимчивости к ОРДС для людей, поскольку на сегодняшний день было идентифицировано более 500 однонуклеотидных полиморфизмов (SNP) Nrf2, а риск ОРДС после тяжелой травмы увеличился у людей с функциональный SNP NRF2 у европейцев и афроамериканцев [90].Для экспериментов, проводимых на животных, гипероксия или LPS-индуцированный ARDS, возможно, являются двумя из наиболее хорошо изученных моделей, которые получают пользу от активации Nrf2, а мыши с дефицитом Nrf2 широко используются в исследованиях, посвященных полезной роли Nrf2 в ARDS. В модели ARDS на грызунах, индуцированной гипероксией, предполагается, что Nrf2 является геном восприимчивости, поскольку функциональные SNP были обнаружены у чувствительных к гипероксии мышей C57BL / 6J (B6) и устойчивых к гипероксии мышей C3H / HeJ (C3), а также у потомства B6C3F2. Сообщается, что у него есть SNP промотора гипероксии [91, 92].Более того, по сравнению с мышами дикого типа у мышей с дефицитом Nrf2 более вероятно развитие ОРДС с повышенной проницаемостью легких, повреждением эпителия и воспалением при стимуляции гипероксией и бутилированным гидрокситолуолом [93, 94]. Недавнее исследование также показало, что специфическая делеция Nrf2 в эпителии дыхательных путей (клетка Клары) в модели индуцированной гипероксией ОРДС достаточна для полного развития острого повреждения легких и замедляет последующее разрешение несоответствующего воспаления легких и эпителиального слущивания [ 95].Для модели LPS-индуцированного ARDS, помимо увеличения восприимчивости и тяжести у мышей с дефицитом Nrf2, в недавнем исследовании также интересно обнаружить, что Nrf2 может оказывать защитное действие на LPS-индуцированный ARDS за счет стимуляции поляризации M2 в макрофагах. используя активатор Nrf2 трет-бутилгидрохинон (tBHQ) и миРНК Nrf2 [96].
Таким образом, в последние годы многочисленные исследования были сосредоточены на защите от ОРДС с помощью активаторов Nrf2, особенно на моделях ОРДС, индуцированных гипероксией или ЛПС.Сообщается, что олеанановый тритерпеноид CDDO-имидазол (CDDO-Im) активирует передачу сигналов Nrf2 / ARE путем нарушения взаимодействий между Keap1 и Nrf2 в цитозоле и оказывает защитные эффекты, такие как ингибирование легочного кровотечения, белкового отека и инфильтрации воспалительных клеток в ОРДС, вызванный гипероксией. Однако эти защитные эффекты почти исчезают у мышей с дефицитом Nrf2 [97]. Кроме того, в другом эксперименте это соединение также подавляло индуцированный аспирацией ОРДС. Важно отметить, что его аналоги, CDDO-метиловые эфиры, уже были использованы в клинических испытаниях для лечения рака (http: // Clinicaltrials.gov), что дает надежду на клиническое применение CDDO-Im. Точно так же витексин и аукубин смягчают воспалительное и окислительное повреждение наряду с подавлением воспалительной передачи сигналов, такой как NLRP3 / NF- κ B, и индукции Nrf2 в модели LPS-индуцированного ARDS в диком типе, но не нокдауна Nrf2. мыши / макрофаги [98, 99]. Наиболее известный индуктор Nrf2, сульфорафан, также оказывает защитное действие на мышиную модель ARDS, индуцированную LPS, путем ингибирования увеличения NF- κ B и активации пути Nrf2, а также, как сообщается, облегчает повреждение легких другой олеиновой кислотой. -индуцированная модель ARDS [100, 101].В самом деле, сообщалось, что большое количество соединений действует аналогичным образом, активируя передачу сигналов Nrf2 в модели ARDS, вызванной LPS, таких как ресвератрол, альфа-липоевая кислота, этилгаллат, кордицепин и сирингин [102–107]. Что еще более интересно, другой мощный активатор Nrf2, tBHQ, защищает от LPS-индуцированного повреждения легких на модели ARDS у мышей, способствуя поляризации макрофагов M2, подавляя поляризацию макрофагов M1 и модулируя баланс между провоспалительными и противовоспалительными факторами [96] .Кроме того, трансфекция Nrf2 также способна усилить эффективность амниотических мезенхимальных стволовых клеток человека (hAMSC) по подавлению воспаления, фиброза и повреждения легких и способствовать дифференцировке hAMSC в клетки альвеолярного эпителия (AT II) типа II с повышенной активностью Nrf2 в LPS-индуцированной модели ARDS у мышей [108].
3.4. Хроническая обструктивная болезнь легких (ХОБЛ)
Хроническая обструктивная болезнь легких (ХОБЛ), распространенная хроническая болезнь легких, характеризующаяся необратимым ограничением воздушного потока, по прогнозам, станет третьей по распространенности причиной смерти к 2020 году [109].Патологическая основа ХОБЛ связана с небольшой обструкцией дыхательных путей и ремоделированием тканей, что может быть результатом аномального воспаления и разрушения паренхимы легких в ответ на различные стимулы, включая генетические / экологические факторы риска и их сложные взаимодействия [110, 111]. Более того, в последние десятилетия оксидативный стресс также был признан важным предрасполагающим фактором, который объясняет патогенез ХОБЛ, а курение сигарет (CS), содержащее богатые оксиданты и другие вредные вещества, долгое время считалось доминирующим экологическим фактором риска развития ХОБЛ. [112].
Воздействие сигаретного дыма может привести к явному окислительному стрессу, воспалению и апоптозу альвеолярных клеток. У здоровых курильщиков, чтобы справиться с такой высокой нагрузкой окислителей, происходит значительное усиление многочисленных антиоксидантных защитных механизмов, среди которых в значительной степени полагается Nrf2. Об этой уверенности может свидетельствовать временная экспрессия Nrf2, индуцированная CS в клетках дыхательных путей человека. Однако наблюдались пониженные уровни Nrf2 и его стабилизатора DJ1 (PARK7) в тканях легких пациентов с ХОБЛ, и многочисленные исследования на людях продемонстрировали, что равновесие NRF2-KEAP1-BACh2 в легочных и альвеолярных макрофагах снижено у пожилых курильщиков. и пациенты с ХОБЛ.Более того, когортное исследование взаимосвязи между полиморфизмом гена Nrf2 и ограничениями воздушного потока у курильщиков также указывает на то, что нарушение Nrf2 может способствовать развитию ХОБЛ из-за чрезмерной нагрузки окислителями и апоптоза в легких [113]. Тяжесть ХОБЛ и возникновение дыхательной недостаточности также могут быть связаны с гаплотипом промотора гена Nrf2, который влияет на его активность [114]. В соответствии с этими данными, мыши с дефицитом Nrf2 более восприимчивы к воздействию сигаретного дыма, у них развивается более тяжелая эмфизема легких и апоптоз, а активность антиоксидантных ферментов подавляется [40].Кроме того, взаимосвязь между Nrf2 и ХОБЛ также исследовалась in vitro: нокдаун Nrf2 увеличивает на 10% экспозицию сигаретным дымом (CSE-) апоптоз, в то время как избыточная экспрессия Nrf2 защищает клетки от апоптоза, вызванного CSE. Примечательно, что делеция Keap1 и последующая активация передачи сигналов Nrf2 в клетках Clara не только защищает клетки от окислительного стресса ex vivo и в эпителиальных клетках человека, но также подавляет окислительный стресс и вызванное CS воспаление in vivo [115]. Кроме того, Nrf2 влияет на острые обострения, связанные с инфекцией, и терапевтические реакции на кортикостероиды при ХОБЛ.Сообщалось, что во время ХОБЛ окислительный стресс нарушает врожденную иммунную защиту и усиливает воспаление. С одной стороны, делеция Nrf2 ухудшает избавление от респираторных инфекций за счет своего воздействия на рецептор-скавенджер макрофагального рецептора с коллагеновой структурой (MARCO). С другой стороны, снижение активности HDAC2 может привести к усилению воспаления и устойчивости к кортикостероидам. Недавнее исследование уже показало, что подвергшиеся воздействию CS и LPS-индуцированные мыши Nrf2 – / – демонстрируют усиленное воспаление легких, которое не может быть облегчено стероидами, и показали, что дефицит Nrf2 может играть важную роль в резистентности к стероидам посредством репрессии HDAC2: рекрутирование HDAC2 важно в опосредовании противовоспалительной активности глюкокортикоидов за счет его взаимодействия с промоторами провоспалительных генов, тогда как дефицит Nrf2 может значительно снизить уровень гистондеацетилазы 2 (HDAC2) и активность деацетилазы [116].
Следовательно, новые методы лечения, такие как активаторы Nrf2, могут оказаться многообещающими для лечения пациентов с ХОБЛ. Некоторые активаторы Nrf2 обладают потенциалом защиты от воздействия сигаретного дыма. Например, при хроническом воздействии CS CDDO-Im индуцирует более значительную активацию Nrf2 и его генов-мишеней, уменьшая вызванный CS оксидативный стресс легких, разрушение ткани и даже легочную гипертензию у мышей дикого типа; однако эти защитные эффекты явно не наблюдаются у мышей с дефицитом Nrf2.Кроме того, интересно, что CDDO-Im не проявляет ингибирующего действия на вызванное CS воспаление, несмотря на то, что он предотвращает развитие эмфиземы, что может свидетельствовать о том, что ингибирования окислительного стресса достаточно, чтобы остановить развитие эмфиземы [117]. Сходным образом, активатор WNT LiCl подавляет эмфизематозные изменения и воспаление легких, индуцированное эластазой или CSE, соответственно, у мышей WT вместе с повышающей регуляцией передачи сигналов Nrf2; однако такие защитные эффекты почти исчезают у мышей с дефицитом Nrf2, зараженных эластазой.Кроме того, защитные эффекты активатора AMPK метформина на стимулированное CSE увеличение воспалительных маркеров в Nrf2-дефицитных клетках NHBE явно не наблюдаются [118]. Более того, два хорошо известных активатора Nrf2, сульфорафан и андрографолид, не только защищают от повреждений, вызванных CS / CSE, но также действуют в борьбе с инфекциями, которые усугубляют ХОБЛ. В недавнем исследовании сообщалось, что сульфорафан противодействует окислительному повреждению, вызванному CSE, с активацией передачи сигналов Nrf2 в линии альвеолярных эпителиальных клеток крысы [119].Кроме того, сульфорафан может усиливать фагоцитоз бактерий (таких как PA и NTHI, выделенные от пациентов с ХОБЛ) альвеолярными макрофагами от пациентов с ХОБЛ, но эта способность отсутствует у макрофагов, трансфицированных миРНК Nrf2, и аналогичные результаты получены для дикого типа и Nrf2. нокаутные мыши. Исследование также показывает, что Nrf2-зависимый бактериальный клиренс сульфорафаном может быть связан с Nrf2-опосредованной регуляцией акцепторного рецептора MARCO, но не зависит от антиоксиданта глутатиона [120].Однако, хотя эти результаты предполагают, что сульфорафан защищает от обострения ХОБЛ и ХОБЛ, введение сульфорафана пациентам с ХОБЛ, к сожалению, не смогло вызвать экспрессию гена-мишени Nrf2 или повлиять на уровни других антиоксидантов или маркеров воспаления в рандомизированном двойном слепом плацебо-контролируемом исследовании. судебное разбирательство в США [121]. Другое типичное соединение для лечения ХОБЛ, андрографолид, представляет собой разновидность лактона, экстрагированного из Andrographis paniculata . Недавние исследования показали, что андрографолид обладает способностью защищать легкие от окислительного повреждения, вызванного сигаретным дымом, и подавлять нетипируемое Haemophilus influenza – (NTHi-) повышенное воспалительное и окислительное повреждение легких в модели мышей с предрасположенностью к CS, которая имитирует обострение ХОБЛ, в то время как механизм действия может быть достигнут за счет индукции Nrf2-опосредованных цитопротекторных ответов [122, 123].Интересно, что это соединение уже применялось в качестве компонента лекарства в некоторых районах Китая. Кроме того, вирус гриппа (FluV) важен при острых обострениях ХОБЛ, и недавнее исследование показало, что мыши Nrf2 – / – , подвергшиеся воздействию CS, показали повышенную смертность и повреждение легких повышенной степени тяжести после инфекции FluV [74]. Следовательно, индукторы Nrf2, которые эффективны при инфекции FluV, такие как куркумин, также могут обеспечивать защиту от некоторых острых обострений ХОБЛ. Действительно, существует множество других соединений, которые, как сообщалось, защищают от повреждения легочной ткани или эпителия, вызванного CS / CSE посредством модуляции Nrf2, такие как ресвератрол, урсоловая кислота, изоформа витамина E , γ, -токотриенол и аспирин- сработал резольвин D1 (AT-RvD1) [124–127].
3.5. Астма
Астма представляет собой сложное респираторное заболевание, характеризующееся хроническим воспалением дыхательных путей, гиперреактивностью дыхательных путей (AHR) и обширной и политропной обратимой обструкцией дыхательных путей [128]. Согласно предыдущим исследованиям, оксидантный стресс играет важную роль в патогенезе астмы у человека, а нагрузка окислителями играет ключевую роль в воспалении дыхательных путей, AHR и даже нечувствительности к стероидам [129, 130]. Как сообщалось, у пациентов с астмой может быть больше проблем с нагрузкой окислителями, чем у здоровых людей, что может быть тесно связано с нарушением активности Nrf2.Недавние исследования также показали, что управляемый Nrf2 GST является возможным маркером предрасположенности к астме у людей: гомозиготный GSTM1-нулевой генотип увеличивает риск астмы, но гомозиготная экспрессия GSTP1 может защищать от астмы [131, 132]. У мышей также сообщалось, что дефицит Nrf2 значительно усиливает оксидативный стресс, вызванный овальбумином или частицами выхлопных газов дизельного топлива (DEP-), воспаление дыхательных путей и AHR в модели астмы [41, 133]. В модели астмы, вызванной OVA, нарушение Nrf2 не только приводит к увеличению уровней эозинофилов, особенно в BALF и тканях легких, у мышей Nrf2 – / – , но также вызывает более высокие уровни нейтрофилов, которые могут быть ответственны за дыхательные пути. ремоделирование при тяжелой астме.В свою очередь, эозинофилы и нейтрофилы производят больше АФК, вызывая повреждение легких. Кроме того, нарушение Nrf2 также может быть связано с более очевидной AHR, гиперплазией бокаловидных клеток, апоптозом эпителиальных клеток и повышенным уровнем цитокинов Th3 в этой модели. В модели астмы, зараженной DEP, мыши Nrf2 – / – также демонстрируют повышенные эозинофилы, AHR, IL-12, IL-13 и тимус, а также регулируемые активацией хемокины (TARC) в BALF. Более того, когда дендритные клетки (DC) как из клеток с дефицитом Nrf2, так и из клеток дикого типа подвергались воздействию твердых частиц (PM), Nrf2 успешно сдерживает выработку проаллергического фенотипа посредством ингибирования окислительного стресса и Th3-направленного проаллергического иммунитета, регулируемого посредством DC [134].Во время этого процесса конститутивная иммуно-поляризующая цитокиновая среда из-за дефицита Nrf2 в DC играет важную роль в увеличивающем стимулирующем эффекте PM на аллергическую сенсибилизацию. Взятые вместе, Nrf2-опосредованные антиоксидантные реакции могут служить важным детерминантом предрасположенности к астме.
В последние десятилетия защитная роль индукторов Nrf2 при астме широко изучается, и овальбумин используется в качестве классического индуктора астмы. До недавнего времени различные исследования изучали защиту сульфорафана от астмы.Введение сульфорафана может подавлять вызванное овальбумином аллергическое воспаление дыхательных путей у мышей, и в недавнем исследовании дополнительно изучалось защитное действие сульфорафана у астматиков. Результаты показывают, что передача сигналов Nrf2 может играть важную роль у людей, чьи бронхопротекторные реакции против метахолина (MCh) улучшаются с помощью сульфорафана [135, 136]. Более того, в модели мышиной астмы, индуцированной Cl 2 , определенной как индуцированная раздражителем астма (IIA) и DEP-стимулированные эпителиальные клетки дыхательных путей, введение сульфорафана с повышенной активностью Nrf2 ингибировало воспаление дыхательных путей [137, 138].Примечательно, что введение сульфорафана здоровым людям не увеличивает экспрессию Nrf2 и его целевого антиоксидантного гена, но защищает от нейтрофильного воспаления дыхательных путей, вызванного озоном [139]. Таким образом, вопрос о том, можно ли использовать сульфорафан в клинической практике, по-прежнему требует дополнительных доказательств из исследований на различных популяциях и аллергенах. Точно так же другой индуктор Nrf2, саппанон A (SA), может защищать от аллергического воспаления дыхательных путей, вызванного OVA, ингибируя рост воспалительных клеток, цитокинов и OVA-специфического IgE в ЖБАЛ и восстанавливая уровень IFN- γ , и механизм действия может происходить из регулируемого Nrf2 баланса Th2 / Th3 [140].Также сообщается, что хорошо известный антиоксидант витамин E ( α -токоферол) защищает от IgE-индуцированной астмы, обращая вспять нарушение активности Nrf2 в альвеолярных макрофагах in vivo и облегчая обострение астмы, стимулированное озоном на мышиной модели, индуцированной OVA. через путь Nrf2, хотя его влияние на симптомы астмы, вызванной OVA, отсутствует [141, 142]. Более того, такие соединения, как витамин D3, форзиазид A (FSA) и основной противомалярийный лекарственный препарат артесунат, все имеют сходные эффекты на уменьшение воспаления дыхательных путей и AHR при астме, индуцированной OVA, посредством активации сигнального пути Nrf2 / HO-1 [143 –145].
3.6. Идиопатический фиброз легких (IPF)
Идиопатический фиброз легких (IPF), важный представитель интерстициального заболевания легких, описывается как хроническое прогрессирующее заболевание легких с фибропролиферацией и избыточным отложением внеклеточного матрикса (ECM), что в конечном итоге приводит к необратимому интерстициальному фиброзу легких. и дыхательная недостаточность [146]. Несмотря на то, что были предприняты большие усилия для борьбы с этим заболеванием, к сожалению, нет сообщений о том, что лечение действительно помогает улучшить выживаемость.Как и многие другие хронические заболевания, патогенез IPF до сих пор неизвестен; однако окислительный стресс тесно связан с предыдущими исследованиями, в которых повышенная окислительная нагрузка уже наблюдалась у пациентов с IPF [147, 148]. Повреждения легочной ткани, вызванные окислительным стрессом, могут иметь разные аспекты. Например, чрезмерное количество ROS приводит к повреждению ДНК и апоптозу в эпителиальных клетках легких и стимулирует прогрессирование IPF за счет запуска активации и высвобождения TGF- β 1, что может ускорять образование ROS, существующее воспаление и рубцевание легких, а также подавлять антиоксидант. экспрессия гена посредством взаимодействия Smad3-ATF3 с Nrf2.Более того, АФК играют важную роль в дифференцировке миофибробластов, которые непосредственно участвуют в патогенезе IPF. Следовательно, неудивительно, что нарушение регуляции Nrf2, главного регулятора оксидативного стресса, как сообщается, вносит значительный вклад в легочный фиброз [149]. В предыдущих исследованиях мыши с дефицитом Nrf2 более восприимчивы к индуктору IPF-подобного фиброза легких и блеомицину, чем мыши дикого типа, и эти мыши с дефицитом более явно проявляют воспаление легких и фиброгенез, а также повышенные индексы фиброза и снижение антиоксидантной реакции [ 38].In vitro siRNA Nrf2 приводит не только к увеличению нагрузки окислителей, но также и к усилению дифференцировки миофибробластов, тогда как нокдаун Keap1 вызывает противоположные эффекты. Тем не менее, доказательства аналогичных выводов на людях еще предстоит изучить, хотя повышенная экспрессия Nrf2, сопровождаемая маркерами оксиданта, обнаруживается в тканях легких пациентов с ИЛФ, особенно когда основной воспалительный ответ и природа оксиданта в общей модели, вызванной блеомицином, все еще контрастируют. с гистологическими особенностями IPF человека.
Несколько активаторов Nrf2 были изучены для защиты от легочного фиброза. Пирфенидон (PFD) является в настоящее время одобренным препаратом для терапии IPF, и его антифиброзная активность в отношении трансформирующего фактора роста- β – (TGF- β -) стимулированных фибробластов и моделей мышей, зараженных блеомицином, связана с восстановлением Равновесие Nrf2 / Bach2 за счет ингибирования Bach2 и активации Nrf2 [150]. Классический активатор Nrf2 сульфорафан также проявляет антифиброзное действие на фибробласты IPF in vitro, обращая характерные признаки миофибробластной дифференцировки (такие как увеличение α -SMA, коллагена I, пролиферация, миграция и сокращение фибробластов) даже при TGF-. β стимуляция, а антифиброзная активность зависит от восстановления окислительно-восстановительного баланса посредством активации Nrf2.Однако в исследованиях сообщалось, что сульфорафан не может защитить от индуцированного блеомицином фиброза легких у мышей, что может быть связано с тем фактом, что эта модель не учитывает эффекты активаторов Nrf2 при фиброзе легких из-за отсутствия активации Nrf2 в фибробластах легких мыши. [151]. Другой индуктор Nrf2, эмодин, аналогичным образом подавлял индуцированные BLM фиброзные повреждения легких у крыс посредством ингибирования гиперпродукции коллагена, эпителиально-мезенхимального перехода (EMT) и экспрессии TGF- β и p-Smad.Кроме того, эмодин может обратить вспять стимулированные рекомбинантным TGF- β 1 ЕМТ-подобные сдвиги в культивируемых альвеолярных эпителиальных клетках [152]. Кроме того, кверцетин значительно индуцирует антиоксидантную защиту посредством активации Nrf2 и подавляет воспаление в клетках BEAS-2B, зараженных блеомицином, а его ингибирующее действие на индуцированный TGF- β фиброз в фибробластах также, по крайней мере, частично опосредовано HO-1, основным нижестоящая цель Nrf2 [153, 154]. Помимо вышеуказанных соединений, антифибротические эффекты берберина и эпигаллокатехин-3-галлата (EGCG) на индуцированный блеомицином фиброз легких на мышиной модели также могут быть, по крайней мере, частично опосредованы активацией передачи сигналов Nrf2, сопровождающейся ингибированием воспаления. и другие биологические события [149, 155].
3,7. Рак легкого
Сообщается, что рак легкого является ведущей причиной смертности от рака во всем мире [156]. При всех типах рака легких немелкоклеточная карцинома легкого (НМРЛ) является наиболее распространенной и была тщательно изучена на ее различных подтипах, в то время как мелкоклеточная карцинома легкого (МРЛ) составляет только примерно 20% рака легких [ 157, 158]. Поскольку это заболевание часто диагностируется поздно, а реакция на существующие лекарства плохая, пациенты с раком легких всегда имеют плохой прогноз.Таким образом, новые возможные лекарственные средства для предотвращения этого заболевания и улучшения результатов по-прежнему остро необходимы.
Чтобы снизить заболеваемость раком, люди пытаются применять химические вещества для детоксикации или удаления канцерогенов [159]. Среди этих потенциальных химикатов индукторы Nrf2 обладают особыми преимуществами в качестве химиопрофилактических агентов [160]. Однако эффекты Nrf2 на патогенез рака легких все еще остаются спорными [161]. Помимо цитопротекторных функций, Nrf2 и его гены-мишени также принимают участие в нескольких онкогенных сигнальных путях, таких как PI3K и K-ras, и связываются с другими факторами транскрипции, структурными белками и эпигенетическими ферментами, участвующими в патогенезе рака [
Biology4Kids .com: Системы животных: Дыхательная система
Ваша дыхательная система – это обмена газов с окружающей средой. Некоторые животные, например земноводные, могут обмениваться газами через влажную кожу. У рыб есть жабры, а у млекопитающих, птиц и рептилий – легкие. Ваша дыхательная система состоит из носа и рта, трубки, называемой глоткой, другой трубки, называемой , трахеи, и ваших легких. У вас два легких, и обмен газов между кровеносной и дыхательной системами происходит в легких.Животным нужно кислорода (O), чтобы выжить. Фактически, всем организмам нужен кислород для завершения процесса сжигания глюкозы в качестве топлива. Даже простейшие и растения нуждаются в кислороде, но по мере того, как вы становитесь более активными, вам нужно больше кислорода. Вот тут-то и важна ваша дыхательная система.
Его цель – доставить кислород в ваше тело. Одним из продуктов клеточного дыхания является углекислый газ. Ваша дыхательная система также помогает организму избавиться от углекислого газа. Хотя у вас есть легкие, у рыб есть жабры, которые служат местом для переноса газов.Какое бы животное вы ни изучали, кислород поглощается, а углекислый газ выходит.
Мы уже упоминали кровеносную систему. С такими газами, как кислород и диоксид углерода (CO 2 ), другие соединения могут попадать в организм через дыхательную систему. Дым может закупорить альвеол в легких. Вы можете вдыхать вирусы и бактерии, которые могут вызвать заболевание. Вы также можете принимать более крупные химические соединения, которые могут отравить ваше тело. Все эти соединения могут попасть в ваш кровоток через дыхательную систему.
Хотя это случается не у всех животных, дыхательная система также взаимодействует с пищеварительной системой. Ваш рот и глотка используются для глотания и дыхания. Есть точка разветвления, где вы найдете надгортанник, который направляет пищу в желудок, а воздух – в легкие. Ваша дыхательная система даже связана с нервной системой в носу, где вы чувствуете запах.
Что может пойти не так с вашей дыхательной системой? Так много всего может пойти не так с этой системой.Он очень уязвим как для токсинов, так и для болезней. Некоторые заболевания, такие как пневмония , могут вызвать наполнение легких жидкостью, и вы больше не сможете получать достаточно кислорода для своего тела.
Курильщики могут накапливать смолу, закупоривая альвеолы в легких и уменьшая количество кислорода, которое может потреблять ваше тело. Экстремальная болезнь, называемая эмфизема , на самом деле разрушает ткани в ваших легких, и ткань не может быть восстановлена. Людям, страдающим эмфиземой, часто приходится дышать чистым кислородом, чтобы выжить.
Healthy Trees – Healthy People (Видео Национальной лесной службы)
Полезные справочные ссылки
Encyclopedia.com:
http://www.encyclopedia.com/topic/respiratory_system.aspx
Википедия:
http://en.wikipedia.org/wiki/Respiratory_system
9000dia Encyclopica Encyclopica
http: //www.britannica.com / EBchecked / topic / 499530 / дыхательная-система человека
Функции, факты, органы и анатомия
Что такое дыхательная система?
Дыхательная система – это сеть органов и тканей, которые помогают вам дышать. Он включает ваши дыхательные пути, легкие и кровеносные сосуды. Мышцы, питающие ваши легкие, также являются частью дыхательной системы. Эти части работают вместе, перемещая кислород по всему телу и очищая отработанные газы, такие как углекислый газ.
Что делает дыхательная система?
Дыхательная система выполняет множество функций. Помимо помощи в вдохе (вдохе) и выдохе (выдохе), он:
- Позволяет говорить и нюхать.
- Доводит воздух до температуры тела и увлажняет его до необходимого уровня влажности.
- Доставляет кислород клеткам вашего тела.
- Удаляет отработанные газы, включая углекислый газ, из организма при выдохе.
- Защищает дыхательные пути от вредных веществ и раздражителей.
Какие части дыхательной системы?
Дыхательная система состоит из множества различных частей, которые работают вместе, чтобы помочь вам дышать. Каждая группа частей состоит из множества отдельных компонентов.
Дыхательные пути доставляют воздух в легкие. Ваши дыхательные пути – сложная система, в которую входят:
- Рот и нос: Отверстия, через которые воздух извне попадает в дыхательную систему.
- Пазухи: Полые места между костями в голове, которые помогают регулировать температуру и влажность вдыхаемого воздуха.
- Глотка (горло): Трубка, по которой воздух доставляется изо рта и носа в трахею (дыхательное горло).
- Трахея: Канал, соединяющий горло и легкие.
- Бронхиальные трубки: Трубки в нижней части трахеи, которые соединяются с каждым легким.
- Легкие: Два органа, которые удаляют кислород из воздуха и передают его в кровь.
Из легких кровь доставляет кислород ко всем вашим органам и другим тканям.
Мышцы и кости помогают перемещать вдыхаемый воздух в легкие и выходить из них. Некоторые кости и мышцы дыхательной системы включают:
- Диафрагма: Мышца, которая помогает легким втягивать воздух и выталкивать его наружу
- Ребра: Кости, которые окружают и защищают ваши легкие и сердце
Когда вы выдыхаете, ваша кровь выносит углекислый газ и другие отходы из организма. Другие компоненты, которые работают с легкими и кровеносными сосудами, включают:
- Альвеолы: Крошечные воздушные мешочки в легких, где происходит обмен кислорода и углекислого газа.
- Bronchioles: Небольшие ветви бронхов, ведущих к альвеолам.
- Капилляры: Кровеносные сосуды в стенках альвеол, которые перемещают кислород и углекислый газ.
- Доли легкого: Разделы легких – три доли правого легкого и две доли левого легкого.
- Плевра: Тонкие мешочки, окружающие каждую долю легкого и отделяющие легкие от грудной стенки.
Некоторые из других компонентов вашей дыхательной системы включают:
- Реснички: Крошечные волоски, которые движутся волнообразным движением, отфильтровывая пыль и другие раздражители из дыхательных путей.
- Надгортанник: Тканевый лоскут на входе в трахею, который закрывается при глотании, чтобы не допустить попадания пищи и жидкости в дыхательные пути.
- Гортань (голосовой ящик): Полый орган, позволяющий говорить и издавать звуки, когда воздух входит и выходит.
Какие условия влияют на дыхательную систему?
Многие состояния могут поражать органы и ткани, составляющие дыхательную систему. Некоторые развиваются из-за раздражителей, которые вы вдыхаете из воздуха, включая вирусы или бактерии, вызывающие инфекцию.Другие возникают в результате болезни или старения.
Состояния, которые могут вызывать воспаление (отек, раздражение и боль) или иным образом влиять на дыхательную систему, включают:
- Аллергия: Вдыхание белков, таких как пыль, плесень и пыльца, может вызвать респираторную аллергию у некоторых людей. Эти белки могут вызывать воспаление дыхательных путей.
- Asthma: Хроническое (длительное) заболевание, астма вызывает воспаление дыхательных путей, которое может затруднить дыхание.
- Инфекция: Инфекции могут вызывать пневмонию (воспаление легких) или бронхит (воспаление бронхов). Общие респираторные инфекции включают грипп (грипп) или простуду.
- Болезнь: Респираторные заболевания включают рак легких и хроническую обструктивную болезнь легких (ХОБЛ). Эти заболевания могут повредить способность дыхательной системы доставлять кислород по всему телу и отфильтровывать отходящие газы.
- Старение: Объем легких уменьшается по мере того, как вы становитесь старше.
- Повреждение: Повреждение дыхательной системы может вызвать проблемы с дыханием.
Как сохранить здоровье дыхательной системы?
Способность выводить слизь из легких и дыхательных путей важна для здоровья дыхательных путей.
Чтобы ваша дыхательная система оставалась здоровой, вам необходимо:
- Избегайте загрязнителей, которые могут повредить дыхательные пути, включая пассивное курение, химические вещества и радон (радиоактивный газ, который может вызвать рак). Наденьте маску, если вы по какой-либо причине подверглись воздействию паров, пыли или других загрязняющих веществ.
- Не курите самостоятельно. Не курите.
- Соблюдайте здоровую диету с большим количеством фруктов и овощей и пейте воду, чтобы избежать обезвоживания
- Регулярно выполняйте физические упражнения, чтобы поддерживать здоровье легких.
- Профилактика инфекций, часто мыть руки и ежегодно делая вакцину от гриппа.
Когда мне следует позвонить поставщику медицинских услуг по поводу проблемы с дыхательной системой?
Обратитесь к своему поставщику, если у вас проблемы с дыханием или боли. Ваш врач послушает вашу грудь, легкие и сердцебиение и будет искать признаки респираторной проблемы, такой как инфекция.
Чтобы убедиться, что ваша дыхательная система работает должным образом, ваш лечащий врач может использовать тесты визуализации, такие как компьютерная томография или МРТ. Эти тесты позволяют вашему поставщику услуг увидеть отек или закупорку в легких и других частях дыхательной системы. Ваш врач также может порекомендовать вам функциональные тесты легких, в том числе спирометрию. Спирометр – это устройство, которое может определить, сколько воздуха вы вдыхаете и выдыхаете.
Обращайтесь к врачу для регулярных осмотров, чтобы предотвратить серьезные респираторные заболевания и заболевания легких.Ранняя диагностика этих проблем может помочь предотвратить их серьезное развитие.
Последний раз проверял медицинский работник Cleveland Clinic 24.01.2020.
Список литературы
Получите полезную, полезную и актуальную информацию о здоровье и благополучии
е Новости
Клиника Кливленда – некоммерческий академический медицинский центр.Реклама на нашем сайте помогает поддерживать нашу миссию. Мы не поддерживаем продукты или услуги, не принадлежащие Cleveland Clinic.
Политика
Дыхательная система
| Зоология для IAS, IFoS и других конкурсных экзаменов
У высших животных основная функция дыхательной системы заключается в транспортировке кислорода из внешней среды в ткани, где он используется для окисления глюкозы с целью производства энергии, а также для переноса углекислого газа, производимого в тканях, и его высвобождения. тела.Кровь вместе с гемоглобином служит для транспортировки газов туда и обратно к местам поглощения кислорода и выделения углекислого газа, которыми могут быть жабры, легкие, щечно-глоточный эпителий, кожа или другие вспомогательные органы дыхания.
Здесь мы изучим анатомию различных типов органов дыхания, которые эволюционировали в серии позвоночных в соответствии с потребностями различных видов и окружающей средой, в которой они живут. Каждое респираторное устройство должно соответствовать следующим основным характеристикам:
- Кровь должна быть отделена от внешней среды – воздуха или воды тонким эпителием.
- Эпителий должен быть проницаемым для диффузии газов через него.
- Респираторный эпителий должен всегда оставаться влажным с пленкой жидкости для осмоса газов.
- Площадь респираторной поверхности должна быть большой, чтобы обеспечить эффективное поглощение кислорода.
- И поток воздуха или воды снаружи, и кровь в капиллярах должны постоянно циркулировать для быстрой замены газов.
GILLS
Жабры у прахордовых
Большой и решетчатый зев у большинства этих животных выполняет двойную функцию: дыхание и улавливание частиц пищи, которые попадают внутрь с потоком воды.Примитивные крыловидные полухордовые (Cephalodiscus и Rhabdopleura) либо не имеют жаберных щелей, либо имеют очень мало щупалец и щупальца, которые помимо сбора пищи также функционируют как эффективные органы дыхания. Balanoglossus обладает большим глотком, имеющим до 700 пар жаберных щелей, что, по-видимому, необходимо в среде обитания норы животного.
Свободноживущие урохордовые, такие как Salpa и Doliolum , не имеют большого количества стигматов или жаберных щелей, поскольку все их тело проницаемо для кислорода, но у малоподвижных асцидий глотка заметно увеличена и перфорирована не менее чем с 200000 стигматов для фильтрующий.
Цефалохордовые используют глотку как для фильтрования, так и для дыхания и, следовательно, несут 150-200 пар жаберных щелей.
Органы дыхания циклостомов
Agnathans имеют 6-15 пар жаберных мешочков, которые являются боковыми продолжениями глотки и содержат внутри жаберные пластинки. Циклостомы называются марсипобранхами, , что означает «жаберные жабры», так как жаберные пластинки расположены в жаберных мешочках. У миксины Myxine всего 6 пар жаберных мешочков, протоки которых соединяются и открываются наружу через одну пару отверстий, тогда как Bdellostoma несет 6-15 пар жаберных мешочков, которые различаются у разных видов и открываются для снаружи самостоятельно.У Myxine позади жаберных мешочков имеется единственный кожно-глоточный проток с левой стороны, который представляет собой модифицированный жаберный мешочек, отводящий лишнюю воду, которая не попадает в жаберные мешочки. Минога, Petromyzon, имеет 8 парных жаберных мешочков зародыша и 7 взрослых особей, которые открываются наружу через независимые отверстия.
Органы дыхания пластиножаберных
У большинства пластиножаберных имеется 5 пар жаберных щелей и пара дыхальцев. Крышка, закрывающая жаберные щели у хрящевых рыб, отсутствует.Демибранх представляет собой пучок жаберных пластинок, прикрепленных с одной стороны от межжаберной перегородки. Таким образом, всего в составе эластожаберных жабер девять пар.
Между двумя полужаберцами лежит межжаберная перегородка, которая поддерживается жаберными хрящами. Кпереди от первой жаберной щели находится дыхальце или псевдожабро. При свободном плавании акул и морских рыб вода обычно поступает через рот.
Кровь к жабрам доставляется пятью парами афферентных жаберных артерий, идущих от брюшной аорты, и, следовательно, они несут дезоксигенированную кровь из сердца.Затем кровь насыщается кислородом в жабрах и собирается петлями четырех пар эфферентных жаберных артерий и переносится к парной дорсальной аорте, две стороны которой встречаются сзади, образуя единую срединную дорсальную аорту, которая снабжает богатой кислородом кровью все тело. .
Жабры костистых рыб
У костистых рыб жабры покрыты жаберной крышкой, состоящей из уплощенных скелетных пластинок, дыхальца нет, как у эластожаберных. Имеется 4 пары жаберных мешочков, каждая из которых содержит по два полужабира, в результате чего общее количество полужаберных у костистых рыб составляет 8 пар или четыре пары полных жабр или голожаберных жабр.
Клеосты всегда дышат с открытым ртом и выбрасывают воду на выдохе, открывая крышку. Жабры у Chondrostei, Holostei и двоякодышащих рыб Neoceratodus демонстрируют частичное сокращение их межжаберных перегородок, что оказывается чем-то вроде промежуточного состояния между эластожаберцами и костистыми мышцами.
НАРУЖНЫЕ ЖАБКИ
Наружные жабры развиваются из наружной стенки глотки или из открытой части жаберной дуги. Они встречаются у личинок миноги, нескольких личинок рыб, Polypterus, двоякодышащих рыб, некоторых личинок костистых рыб и всех личинок и некоторых взрослых особей амфибий.У хондростейных костистых рыб есть единственная пара личиночных жабр, Polypterus, , имеющая длинную ось, несущую жаберные пластинки.
Африканские и южноамериканские легкие рыбы обладают 4 парами перистых наружных жабр. Личиночные формы некоторых земноводных и некоторых взрослых уродел имеют наружные жабры, которые возникают просто как складки кожи на поверхности III, IV и V жаберных дуг, но слабо поддерживаются скелетной системой. Perennibranch земноводных, как Necturus и Proteus , сохраняют внешние жабры на протяжении всей жизни вместе с 2 или 3 парами жаберных щелей, которые не функционируют, поскольку вода не проходит через глотку.
Вместо этого в воде жабры раскачиваются с помощью мышц, прикрепленных к основанию оси жабр для дыхания. Личинки амфибии без конечностей, Caecilia, имеют пару исключительно крупных листовидных жабр с обильным кровоснабжением. Саламандры, населяющие горные ручьи, например Eurycea и Salamandrina, , которые принадлежат к семейству Plethodontidae, не имеют ни жабр, ни легких для дыхания и выживают только при кожном дыхании.
ВОЗДУШНЫЙ ПЕРЕХОДНИК
За исключением бесчеловечных, хрящевых рыб и немногочисленных донных костистых рыб, все рыбы несут заполненный газом воздушный пузырь на дорсальной стороне кишечника, который служит гидростатическим органом.На брюшной стороне мочевого пузыря находится сильно васкуляризованная область, называемая красной железой , которая снабжается кишечной артерией и воротной веной и которая обладает уникальной способностью извлекать свободный кислород из крови и выпускать его в воздушный пузырь, чтобы его надуть.
Небольшой дивертикул в виде мешочка, называемый овальной формы , который может закрываться или открываться мышцами сфинктера, является местом реабсорбции газов. Секреция и абсорбция газов в плавательном пузыре происходит под контролем вегетативной нервной системы в зависимости от глубины, на которой плавает рыба.
У Cypriniformes (Teleostei) – серия из четырех мелких костей (трипус, интеркаларий, скафий и claustrum), , происходящие от первых трех позвонков и называемые Weberian Косточки , соединяют передний конец воздушного пузыря с непроницаемой пазухой перепончатого лабиринта. Звуковые колебания, получаемые воздушным пузырем от окружающей воды, передаются во внутреннее ухо через это уникальное устройство, чтобы дать этим рыбам некоторую способность слышать.
У некоторых рыб, например ганоидов, карпов и сомов, пневматический канал соединяет воздушный пузырь с пищеводом. Такое состояние называется физостомоз (гр. физо = мешок; стома = отверстий). Рыб, у которых нет такого пневматического канала, соединяющего воздушный пузырь, называют Physoclistous (Gr. Physo = мешок; clista = закрытый).
Сравнительное исследование воздушных пузырей у разных групп рыб и поразительное сходство плавательного пузыря и легкого предполагают филогенетическую связь между ними.Принято считать, что легкие произошли от воздушного пузыря рыб. Однако недавние данные свидетельствуют об обратном: легкие сначала развивались у рыб для пополнения запасов кислорода из воздуха, а затем они превратились в плавательный пузырь по мере увеличения концентрации кислорода в воде.