

Тарань и вобла отличия: Страница не найдена | sundaria.su
Лучшее пиво мира на BeerMonsters.ru » Blog Archive » Таранка или вобла
Если вы живете в Париже – вы, конечно парижанин, ну или парижанка. А если в Москве – соответственно москвич (москвичка). И при всем при этом, где бы вы ни проживали, вы остаетесь человеком, мужчиной или женщиной, блондином или брюнеткой, с карими или зелеными глазами. Точно так же дело обстоит и с рыбой из семейства карповых, рода плотва (Rutilus rutilus): морская плотва, «проживающая» поблизости от устьев рек, впадающих в Черное и Азовское моря именуется таранью (лат. Rutilus heckelii), а та же морская плотва, «прописанная» неподалеку от устьев рек, впадающих в море Каспийское – это уже вобла (лат. Rutilus caspicus).
«Неподалеку» и «поблизости» – от того, что метать икру по весне и тарань, и вобла отправляются в эти самые устья, в тишину и покой камышовых зарослей и мелководий. А размножившись, худющие и оголодавшие рыбины возвращаются в море, где для них, плотоядных, обожающих малоподвижных беспозвоночных (всяких моллюсков и ракообразных), самое раздолье. За такие метания тарань и вобла называют плотвой полупроходной, в отличие от плотвы обычной, жилой, промышляющей личинками насекомых и растениями в прудах, реках и озерах.
Это различие в рационах и образе жизни сказалось, конечно, не только на названии: более жирная и мясистая, довольно интенсивно набирающая в весе тарань и вобла всегда была и более привлекательна для желающих полакомится рыбкой. А путешествия ее на нерест плотными косяками в доступных для лова местах превратили тарань и воблу в объект отличного и беспроигрышного промысла. Выловленные горы воблы (или тарани) перерабатывались самым древним и незамысловатым (но при этом самым надежным и незатратным) способом: ее на несколько суток непотрошеной и живой отправляли в крепкий соляной раствор (тузлук), после чего провяливали (просто на воздухе) порядка недели.
В результате получался непередаваемо вкусный, насыщенный и недорогой полуфабрикат, из которого наши далекие предки беспрепятственно готовили самые разнообразные горячие блюда, или же с удовольствием наминали просто так – под домашнее пиво. И сей пищевой продукт отнюдь не входил в число деликатесов: по крайней мере, до революции промысел воблы и тарани предполагал исчисление этой рыбы не штуками, килограммами или тоннами, а горами и возами. Однако со временем размеры эти существенно уменьшились, и, в конце концов, под таранкой (или таранькой) стала подразумеваться не только тарань или вобла, но и многие другие сорта вяленой и сушеной рыбы.
И сей пищевой продукт отнюдь не входил в число деликатесов: по крайней мере, до революции промысел воблы и тарани предполагал исчисление этой рыбы не штуками, килограммами или тоннами, а горами и возами. Однако со временем размеры эти существенно уменьшились, и, в конце концов, под таранкой (или таранькой) стала подразумеваться не только тарань или вобла, но и многие другие сорта вяленой и сушеной рыбы.
К 60-м – 70-м годам прошлого века вобла и таранька (готовая к употреблению тарань) стали настолько дефицитны, что перешли в разряд деликатесов, и вдалеке от ареалов обитания их можно было уже встретить разве что в составе «продуктовых заказов», которыми наши особо везучие сограждане одаривались на предприятиях по большим праздникам, или же в валютных магазинах «Березка». Да-да, валютных, а что вы думаете? Для многих иностранцев (а особенно – из числа наших бывших соотечественников) хорошо приготовленная вобла – это деликатес, вполне сравнимый с икрой красной и икрой черной, поскольку такой способ приготовления, также как и такие сорта рыбы, не воспроизводятся больше ни в одной стране мира.
Какой же должна быть качественно приготовленная таранка (или таранька), чтобы ею не стыдно было закусить отменное пиво? Ну, в первую очередь, как мы уже с вами условились, она должна быть приготовлена именно из тарани или воблы. Далее – обращаем внимание на запах: аромат у нужной нам таранки изумительно чистый, прозрачный, и отчасти даже содержащий в своем составе нотки свежих огородных огурцов. Никакой затхлости, горечи, и уж тем более кислятины. В общем, спутать его с чем-то иным, так же как и сравнить – довольно проблематично. Далее пробуем рыбину на изгиб, вспоминая основы сопромата. Искусно высушенная вобла непременно гибкая и упругая, пружинистая, но не гуттаперчевая. Она способна согнуться сплошным листом, но не до конца: если вы можете притянуть с лёгкостью голову к хвосту, таранька либо не досушена, либо отсырела. Ну а если вобла представляет собой сплошной монолит, который, дабы облегчить ее расковыривание, предварительно придется отбить об надежный стол (как во множестве советских фильмов) – она безнадежно пересушена.
Ну а если вобла представляет собой сплошной монолит, который, дабы облегчить ее расковыривание, предварительно придется отбить об надежный стол (как во множестве советских фильмов) – она безнадежно пересушена.
После проверки на прочность подвергаем приглянувшуюся нам рыбку визуальному осмотру: у правильно приготовленной воблы или тарани сквозь спинку должно просвечивать солнце. Если рыба прошла все этапы вашего тестирования на «отлично» — даже не сомневайтесь: вас ожидает незабываемый карнавал вкусовых и эмоциональных ощущений: после непродолжительной борьбы с ее одеяниями взору откроется янтарная, поблескивающая нежным жирком мякоть, которая, кажется, сама излучает теплый золотистый свет, приглашая вас насладится нежной, чуть вязковатой упругостью, в меру соленой, чуть терпковатой, с ароматом морских странствий и вольного ветра. Ну а если совсем повезет – внутренности вашей красавицы будут битком набиты золотистой икрой, вкуснее которой даже невозможно придумать что бывает еще.
Конечно, наслаждаться такой роскошью без пива – это значит просто упускать из события все самые ключевые моменты. Пусть даже безалкогольное, пусть в минимальном количестве – но пиво отменного качества должно на столе, украшенном воблой или таранькой, присутствовать непременно. В идеале, конечно, это пилзнеры, и желательно – «Пилзнер Праздрой» ибо никакая другая марка своей хмелевой горчинкой не подчеркнет так изящно едва ощутимую горчинку воблы, ни какая другая плотность не способна соперничать с плотностью и сочащимся жирком таранки, и никакой другой цвет напитка так идеально не сочетается своей янтарностью с янтарным светом, источаемым икрой настоящей воблы или тарани.
Ну а если пиво подкачало – не унывайте. Во-первых, настоящая вобла или таранька тем и славятся, что готовы ждать вас годами – в плотно закрытой капроновой крышкой банке, в прохладном темном месте они безропотно потерпят, пока вы соберете для них достойных компаньонов. Другой вопрос – сможете ли ждать вы, зная, что где-то совсем по близости, на расстоянии вытянутой руки находится счастье. Если силы воли отложить праздник живота не хватает – наплевав на все, сосредоточьтесь на рыбке. А посредственное пиво – что ж, знающие люди утверждают, что им очень успешно запах воблы или таранки уничтожается, если ополоснуть им в конце трапезы руки…
Если силы воли отложить праздник живота не хватает – наплевав на все, сосредоточьтесь на рыбке. А посредственное пиво – что ж, знающие люди утверждают, что им очень успешно запах воблы или таранки уничтожается, если ополоснуть им в конце трапезы руки…
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Метки: вобла, закусь, рыба, таранка
Google+
ᐉ Тарань – ✅ ribalka-snasti.ru
Тарань (лат. Rutilus heckelii ) — представляет разновидность обыкновенной плотвы (Rutilus rutilus). Как отличительные признаки по сравнению с обыкновенной плотвой указывается большая высота тела, чешуя меньшего размера, меньшее число лучей в заднепроходном плавнике (именно 9-10), более толстые зубы и черноватые края парных плавников. Длина обыкновенно 25-35 см, вес часто до 1,8 кг. Водится в морях Чёрном и Азовском, входя в реки для метания икры весною (ход может начинаться уже зимою) и отчасти осенью на зимовку. Большая часть зимует, по-видимому, в море у устьев рек. Вообще Тарань не поднимается в реки высоко. Нерест происходит в конце марта или начале апреля в камышах и траве, и по окончании его тарань уходит в море.
Латинское название: Rutilus rutilus heckeli
Другие названия: таранка
Семейство: Карповые
Род: Плотва
Тип: полупроходная
Образ жизни: пелагический
Тип питания: мирный
Ареал обитания: бассейн Черного моря
Тарань – лучеперая рыба, представитель семейства карповых. От родственницы плотвы не сильно то и отличается. У тарани более высокое, распластанное тело, чешуйки гораздо мельче, зубы мощнее и толще. Туловище тарани вытянутое, высокое, прижатое с боков.
Самцы и самки окрашены одинаково – серебристо-металлический тон. Верхняя часть головы и поверхность спинки имеют буровато-оранжевый оттенок, брюшко светлое. Боковая линия слабо выражена. Голова небольшая, треугольная. Рот маленький, нижнерасположенный, глаза золотистые, крупные. Плавники имеют зеленовато-рыжий оттенок и темный, почти черный тонкий ареал по краям. Максимальный размер тарани – до 35 см в длину при весе до 1,5 кг. Но в основном встречаются особи по – 300-400 гр.
Голова небольшая, треугольная. Рот маленький, нижнерасположенный, глаза золотистые, крупные. Плавники имеют зеленовато-рыжий оттенок и темный, почти черный тонкий ареал по краям. Максимальный размер тарани – до 35 см в длину при весе до 1,5 кг. Но в основном встречаются особи по – 300-400 гр.
Места обитания
Ареал обитания тарани это реки азовско-черноморского бассейна. Как и вобла, она бывает двух видов «проходная» и «туводная». Предпочтение тарань отдает прибрежным зонам, так как вода в этих местах теплее, да и корм найти значительно проще. На икромет она стремиться зайти вверх по течению рек, а после него возвращается на обжитые места. Не любит тарань быстрины. Ее больше привлекают притоки и затоны или неглубокие ямы, где течение заметно слабее.
Тарань в отличие от карася и линя не любит илистого дна, предпочтение она отдает песчаному дну. В зимнее время она уходит на большие глубины, а на мелководье выходит только в редкие периоды кормежки, хотя в спячку, подобно сазану не впадает.
Проходная тарань подходит к устьям рек уже с середины февраля, а с наступлением первых оттепелей устремляется к местам нереста.
Образ жизни тарани с ее постоянными миграциями располагает к созданию больших стай. Передвигаться из моря в реку на нерест безопаснее в большой компании. Летом и осенью она кормится здоровой морской пищей, которую находит здесь в изобилии. Осенью происходит второй ход рыбы к местам зимовки. Тут уже тарань, подобно вобле , образует огромные косяки, которые устремляются в устья рек.
Тарань относят к эврифагам, то есть к всеядным рыбам, а это значит, что она может питаться и растительной и животной пищей. Весной, попадая в реки с буйной растительностью, она употребляет в пищу нитчатые водоросли, которые растут на небольшом течении, оплетая камни или сваи. Этот вид корма рыбаки называют «шелковником». Любит водоросли и высшую растительность жилой вид тарани.
После икромета тарань пытается восполнить силы и переходит на животный корм, основу которого составляют моллюски, разнообразные беспозвоночные и их личинки. Полупроходная тарань поедает двустворчатых моллюсков, таких как дрейсену, адакну и монодакну. Не отказывается она и от различных ракообразных, например, бокоплавов и мизид.
Полупроходная тарань поедает двустворчатых моллюсков, таких как дрейсену, адакну и монодакну. Не отказывается она и от различных ракообразных, например, бокоплавов и мизид.
Крупные особи могут позволить себе полакомиться и мальками, если их появляется много. Ближе к зиме, тарань, возвратившись в устья рек, бродит в поисках пищи. Основу ее рациона в зимнее время составляет мотыль и личинки различных насекомых.
Размножение
Как и у большинства рыб, нерест тарани происходит весной. Обычно этот период приходится на март — май. Основным показателем для нереста является температура воды. Она должна быть не холоднее +8 градусов. Огромными косяками проходной вид этой рыбы устремляется в реки, но предпочитает не подниматься высоко. Самки достигают половой зрелости на третьем году жизни. Во время нереста меняется окраска тарани. Она становится значительно интенсивнее. На теле крупных самок и самцов появляются характерные бугорки.
Самки мечут икру на водную растительность, которая служит одновременно и защитой для нее. Отложение икры у тарани происходит единовременно. Икринки диаметром 1,5 мм приклеиваются к стеблям и листьям растений. Для развития икры требуется от 9 до 14 дней, после чего из икры вылупляются личинки размером до 6 мм. Сначала молодь питается водорослями, он очень быстро переходит на употребление мелких беспозвоночных. Молодь полупроходного вида растет быстрее свих жилых собратьев.
С мест нереста мальки тарани пассивно плывут по течению, но спустя небольшой промежуток времени их активность возрастает и они устремляются в море. Плодовитость полупроходного вида тарани так же значительно выше. Если самка жилого вида откладывает одновременно до 100 икринок, то у самок полупроходного вида их количество возрастает вдвое.
Сезоны ловли
Весной огромные косяки тарани плывут к местам своего нереста. Кажется, как бы ни сейчас праздновать начало рыбацкого сезона. В это время таранью можно просто уловиться. Однако, именно в это время ловля тарани в некоторых регионах запрещена даже на любительские снасти. Поэтому, если вы не хотите иметь проблемы с законом, разумнее будет подождать начала лета.
Однако, именно в это время ловля тарани в некоторых регионах запрещена даже на любительские снасти. Поэтому, если вы не хотите иметь проблемы с законом, разумнее будет подождать начала лета.
В это время клев этой рыбы становится интенсивным, поскольку после нереста тарань становится удивительно прожорливой. Она возвращается к местам своего постоянного проживания, по пути, заглатывая все, что попадается ей на глаза.
Осенняя рыбалка на тарань тоже может порадовать рыболовов, если правильно выбрать место. Жилой вид этой рыбы можно поискать на глубоководных участках рек, а проходной вид в их устьях и в море.
Ближе к зиме морская тарань совершает второй переход в реки. Она стремится нагулять жир перед длительной зимовкой, следовательно, клев в это время может быть интенсивным. С первым ледоставом открывается зимний сезон ловли этой рыбы. Она не впадает в зимнюю спячку и охотно клюет на протяжении всей зимы.
Способы ловли
В Азово-Кубанском районе тарань не только самая популярная, но и одна из самых ценных рыб. Поэтому в период нереста (весной) – промышленный и любительский отлов тарани категорически запрещен. В остальное время её ловят в достаточно больших количествах, для засолки и вяления.
Лучшим периодом для ловли тарани является лето, когда рыба вернувшись с мест нереста и изголодавшись, с жадностью набрасывается на любую наживку – от рачьего мяса до хлебного мякиша.
Но лучшими остаются все таки навозный червь и опарыш. Снасть может быть в принципе любая. Начиная от обыкновенной ( маховой ) поплавочной удочки и различными видами зимних удочек, заканчивая фидером, который будет просто незаменим.
Видео: Тарань
Рыба тарань азовско-черноморская (Rutilus heckelii) представляет собой по сути одну из разновидностей обыкновенной плотвы. Однако отличительными признаками тарани по сравнению с плотвой считается большая высота тела рыбы, меньших размеров чешуя, меньшее количество лучей в плавнике заднепроходном, черноватая краевая линия парных плавников и утолщенные зубы.
Средняя длина рыбы тарань достигает 35 сантиметров при максимальном весе в 1.8 килограммов. Как правило, тарань водится в Черном море и Азовском, для метания икры весною входя в реки и частично осенью для зимовки. Большая часть особей тарани зимует в морях у устьев рек, но вообще эта рыба в реки высоко не поднимается.
Из-за того, что рыба тарань поступала в торговую сеть, как правило, в вяленом виде, представляя собой излюбленную пищу народов Дона, Кубани и всего Приазовья, под названием тарань сегодня понимают товар, получаемый не только из данного вида рыб, но и при смешивании разных мелких рыб. Кстати, на Дону называют таранью и привозимую в изобилии с Волги вяленую воблу.
Раньше Азовское море наряду с впадающими в него реками, особенно Доном, давали поистине колоссальные количества рыбы тарань, которая составляла немаловажный предмет рыбного промысла. Так, целые горы сушеной тарани привозили на далеко славившийся в прежние времена рыбный рынок, который находился на Гниловской станице, а откуда рыба развозилась чумаками по всей Украине.
На сегодняшний день наблюдается полное оскудение Азовского бассейна рыбой тарань, которую выловить более негде. Причиной такого исчезновения можно назвать не только безграничный лов рыбы тарань до ее нереста, а также уничтожение молодых особей. Неблагоприятные стихийные обстоятельства для развития выметанной икры оказывают также важное влияние на создание печального положения дел с рыбой тарань в современное время.
Польза тарани
Как известно, содержание тех или иных полезных веществ в составе рыбе может изменяться в зависимости от ее возраста, условий жизни и питания, сезонных изменений и даже пола. Относительно пользы тарани можно сказать, что этот вид рыбы имеет в своем составе немало легкоусваиваемого белка, что крайне благоприятно сказывается на здоровье человека.
Кроме того, польза тарани для организма человека заключается и в наличии некоторых витаминов (в частности РР), а также минеральных солей.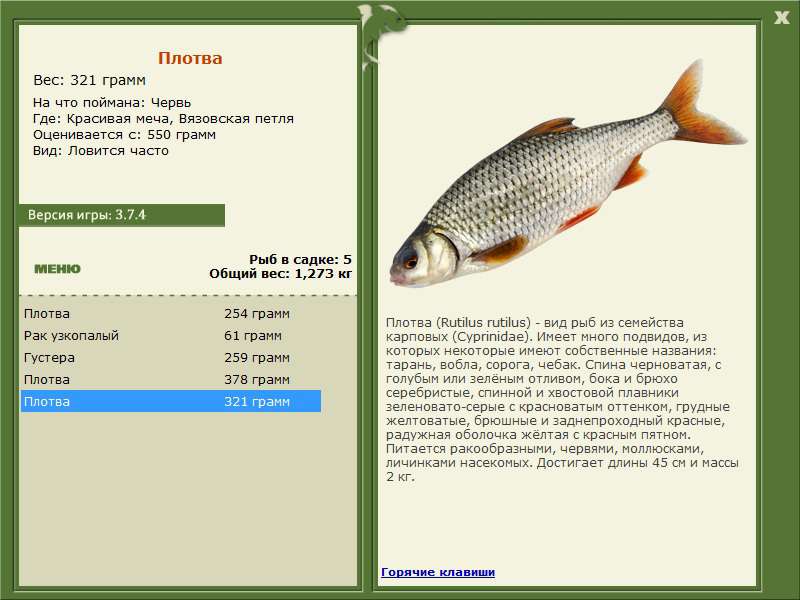 Среди последних стоит отметить серу, хлор, молибден, никель, цинк, хром и фтор. Калорийность тарани в свежем виде составляет около 88 ккал.
Среди последних стоит отметить серу, хлор, молибден, никель, цинк, хром и фтор. Калорийность тарани в свежем виде составляет около 88 ккал.
Что за рыбы вобла и тарань
Рыба тарань и вобла – представители семейства карповых. Они являются полупроходными подвидами обыкновенной плотвы. Для простого обывателя разницы между ними нет, но для опытного рыболова эти подводные жители отличаются. Об этом и пойдет разговор ниже.
Разновидности плотвы
Многие ихтиологи и опытные рыболовы выделяют несколько подвидов плотвы. Среди них есть полностью пресноводные представители, есть полупроходные рыбы, либо проживающие в солоноватой воде:
- плотва обыкновенная – населяет все пресноводные водоемы умеренных широт;
- плотва сибирская, чаще называемая чебак – проживает на Урале, Дальнем Востоке, в Сибири;
- аральская плотва – обитает в Аральском море и его притоках;
- каспийская вобла – водится в Каспийском море, Балхашском озере и низовьях Волги;
- азовско-черноморская тарань – живет в Черном и Азовском море, низовьях Днепра, Дона и Дуная.
Наибольшее промысловое значение имеет вобла и тарань. Плотва обыкновенная и чебак являются преимущественно объектом любительского лова. Хотя на первых также охотятся и рыболовы, особенно в период их весеннего хода, когда рыба идет на нерест.
Рыба вобла – подвид плотвы, населяющий Каспийское море. Такие подводные жители называются эндемиками, поскольку они живут на относительно ограниченной акватории и являются ее неотъемлемой составляющей. В течение года нагуливается на открытых просторах, а весной мигрирует на нерест в большие притоки, в первую очередь Волгу.
Отличие воблы от плотвы заключаются в более крупных размерах. Благодаря хорошей кормовой базе, рыба достигает внушительного веса. Среднестатистическая масса пойманных особей составляет порядка 300 граммов, нередки поимки экземпляров свыше килограмма.
Кроме этого, вобла имеет отличия по некоторым второстепенным морфологическим признакам:
- плавники имеют темный цвет с выраженной черной оторочкой;
- глаза серебристого цвета;
- над зрачками черные пятна.

Вобла зимует в море. В начале весны, иногда еще подо льдом, она начинает движение в реку. Далеко вверх не поднимается. Заходит в различные ерики, протоки, старицы и рукава, где в камыше откладывает икру.
Отнерестившись, вобла разрозненными косяками скатывается обратно в море. Здесь рыба нагуливает вес, активно питаясь. Такое поведение длится до поздней осени. Затем каспийская плотва сбивается в косяки, подходит ближе к устьям рек, где проводит зиму.
Тарань – полупроходная форма плотвы, населяющая Черной и Азовское море. Встречается в лиманах и заливах, устьях больших рек, таких как Дон, Днепр, Кубань, Дунай, Миус и другие, в периоды сезонной миграции заходит в них, но высоко не поднимается.
Рассмотрим, чем тарань отличается от плотвы:
- покрыта чешуйками меньшего размера;
- имеет более высокое тело;
- парные плавники имеют черноватые края;
- более толстые губы;
- меньшее число лучей на анальном плавнике.
Тарань и плотва имеет разницу и в размерах. Благодаря качественной кормовой базе, первая вырастает до 50 см при массе тела до 2 кг. Тогда как пресноводная представительница ихтиофауны редко вырастает до 1–1,2 кг, и то, подобные экземпляры встречаются на водохранилищах и в больших реках.
Азовско-черноморская тарань плотва с апреля по сентябрь нагуливается в открытом море. Рыба активно отъедается, набирая вес перед холодным периодом. В октябре, при понижении температуры воды до 15 градусов, плотными косяками подходит в прибрежную зону, обосновывается в лиманах и устьях рек. Здесь и проводит зиму.
В начале весны можно наблюдать преднерестовый ход тарани. Рыба плотными косяками двигается вверх по течению, заходя в старицы ерики и протоки, на мелководья и спокойные участки. Мечет икру в утренние и вечерние часы около прошлогодней водной растительности.
Фото 1. Плотина — уловистое плотвиное место во время нерестового хода.
Выклюнувшийся малек питается зоопланктоном:
Подросшая молодь, скатившись в море, переходит на питание червями, моллюсками, ракообразными.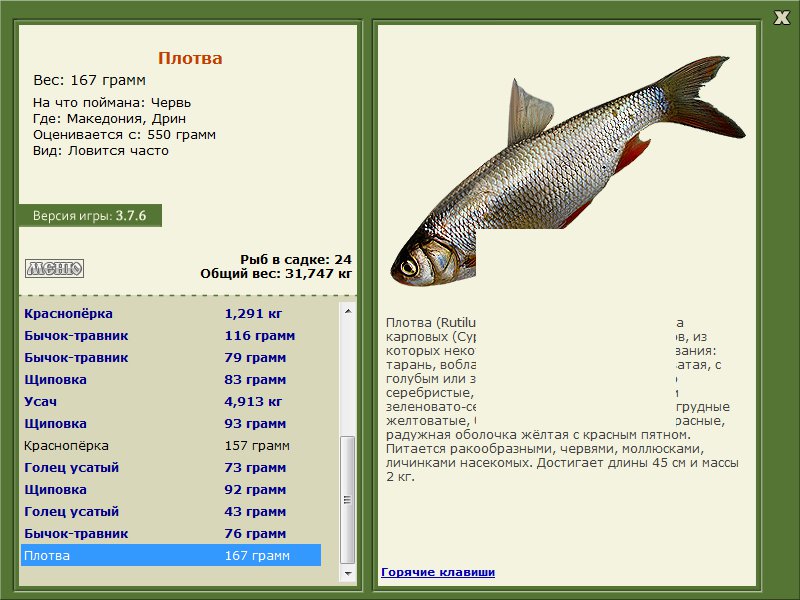
Отличия между воблой и таранью
Основным отличием воблы от тарани является ареал их обитания. У первой он более ограничен – Каспий и низовья Волги. У второй представительницы ихтиофауны – Черное и Азовское море, устья больших рек, впадающих в них, и их нижнее течение.
По внешнему виду рыбы очень похожи. Обе имеют более высокое тело, чем у обыкновенной плотвы. Достигают внушительных размеров, но есть и некоторые различия:
- у тарани чешуя мельче;
- вобла имеет серые плавники, тарань – с красноватым отливом;
- у тарани рот полунижний с толстыми губами, у воблы – ближе к прямому, как у плотвы обыкновенной.
Оба подвида комфортно чувствуют себя и в подсоленной, и в пресной воде. Склонны группироваться многочисленными косяками, состоящими из особей приблизительно одного размера.
Ловля воблы и тарани
Ловля тарани и воблы особо не отличается между собой. За этими рыбами лучше всего отправляться в весенний период, когда у них наблюдается преднерестовый ход. В этом время ужение данных подвидов плотвы наиболее результативно. Также на них успешно охотятся после икромета и во второй половине осени.
Тарань и вобла – стайные рыбы, которые перемещаются по акватории. Потому основной задачей для рыболова является обнаружение их косяков, а остальное дело техники – грамотно подобранных и настроенных снастей, выбранной приманки и прикормочной смеси.
Ловля тарани и воблы ведется поплавочными и донными снастями. Последние более востребованы, разнообразны и результативны. Это может быть обычный фидер или пикер, донка либо грузовка, резинка либо закидушка. Базовая снасть на основе удилища и катушки выглядит следующим образом:
- Удилище фидерное либо для донной ловли длиной от 2,1 до 4,2 метра, в зависимости от дистанции и условий ужения. Тест по приманкам от 100 до 200 грамм.
- Катушка безынерционная силовая с передаточным числом порядка 4,2:1, размером 4000–6000. Представители «старой школы» ловят инерционками советского образца.

- Монофильная леска толщиной от 0,3 до 0,4 мм либо плетеный шнур с разрывной нагрузкой до 5–7 кг.
К основной леске привязывается поводок из мононитки толщиной 0,2–0,25 мм длиной от 30 до 80 см, в зависимости от активности рыбы и горизонта, в котором она поднимается вверх по течению. Используется кормушка либо свинцовый груз нужного веса. Крючок подбирается под приманку и потенциальный размер добычи.
Фото 2. Ловля плотвы на фидер.
Если у рыболова нет подходящего удилища, то выручит простейшая снасть на тарань, выполненная в виде банальной закидушки:
- Берем монофильную леску толщиной 0,35–0,4 мм длиной около 100 метров и наматываем ее на мотовило.
- На конце лески привязываем массивное свинцовое грузило, необходимое для удержания оснастки в нужной точке.
- На основной леске делаем несколько петель длиной 2–3 см для крепления поводков. Первую вяжем в 10–15 см от груза. Расстояние между ними составляет 20–40 см.
- Способом «петля в петлю» крепим поводки из лески диаметром 0,22–0,26 мм длиной до 30см.
- Остается к поводкам привязать одинарные крючки подходящего размера.
Представленная снасть хоть и уступает в чувствительности фидеру и пикеру, но имеет преимущественно в простоте изготовления и минимальным затратам на покупку ее элементов.
Ловля тарани и воблы на поплавочную удочку практикуется в местах, где не требуется дальний заброс, а ужение ведется накоротке. Это могут быть узкие протоки, ответвления, протяженные канавки под берегом, мелководные участки с прошлогодней водной растительностью.
Поплавочные снасти для ловли тарани и воблы в этом случае подбираются легкие, но достаточно надежные, чтобы вываживать увесистую рыбу. Удилище используется телескопическое, длиной в 5–6 метров. Леска монофильная, диаметром до 0,3 мм с разрывной нагрузкой не менее 3–4 кг.
Некоторые рыболовы ловлю воблы и тарани осуществляют матчевыми удочками. При этом используют безынерционные катушки размером 1500–2000 и более тонкие лески, по сравнению с донными снастями. Это возможно в местах с умеренным течением, где глубины колеблются от 1 до 3 метров.
Это возможно в местах с умеренным течением, где глубины колеблются от 1 до 3 метров.
Вобла, тарань, как и плотва, относятся к мирным подводным жителям, которые ловятся на приманки животного и растительного происхождения. Из наживок для их ужения можно отметить:
- червя;
- мотыля;
- опарыша;
- личинку ручейника;
- личинку репейника;
- кузнечика.
В холодной воде самыми предпочтительными приманками являются червь и мотыль. По мере потепления начинают работать другие перечисленные наживки.
В теплое время года, особенно в жаркие летние месяцы, рыба отдает предпочтение растительным приманкам. Из самых ходовых и доступных насадок для ловли тарани и воблы можно рекомендовать:
- пропаренную пшеницу;
- консервированную кукурузу;
- тесто.
Фото 3. Для сохранности кукурузу заливают подсолнечным маслом.
Некоторые рыболовы успешно применяют мелкие ароматизированные бойлы, банальный хлебный мякиш, консервированный или свежий горошек.
Нельзя забывать про такую приманку для плотвы, как пенопласт. Являясь несъедобным, он привлекает рыбу за счет своей положительной плавучести и колебаний на течении. Шарики «пены» актуальны в донных оснастках, когда основную насадку или наживку надо поднять выше, поскольку тарань кормится в толще воды.
Ловля тарани и воблы в их преднерестовый ход не требует использования прикормки, поскольку рыба непрерывно движется на места икромета, и нет смысла пытаться удерживать ее в заданной точке. Гораздо эффективнее активно искать рыбу и рыбачить непосредственно на ее «тропах».
В теплую пору рекомендуется применять прикормочную смесь, которая позволит привлечь рыбу в зону ужения и удержать ее там определенное время. Для этого можно использовать фабричные «сыпучки», а можно приготовить недорогую, но качественную прикормку своими руками. Делается это так:
- В обычную эмалированную кастрюлю надо налить полтора литра холодной воды.
- Добавляем в нее 300–500 граммов гороха, предварительно залитого водой с ночи.
- Доводим воду до кипения, убавляем огонь, накрываем кастрюлю крышкой и томим горох около часа.
- Сваренный горох перетираем до состояния пюре. Делается это с помощью мясорубки, блендера или других подручных средств.
- В подготовленное гороховое пюре добавляем столовую ложку подсолнечного масла.
Вместо растительного масла горох можно перетереть с очищенными семечками подсолнечника.
Приехав на рыбалку, подготовленное пюре смешиваем с грунтом, добытым на берегу либо на дне водоема. Формируем шары диаметром от 5 до 10 см. Сперва делаем стартовый закорм, забросив их в воду от 7 до 10 штук. После подбрасываем по несколько штук через 15–30 минут.
Промысловая ценность
Тарань и вобла представляют высокую промысловую ценность. Заготовкой этих видов рыб занимаются в весенний период, когда наблюдается их массовый нерестовый ход в низовьях больших рек. В остальное время целенаправленно за ними не охотятся.
Вобла и тарань хороша в следующем виде:
Промысловая ловля тарани и воблы в последнее время стала жестче контролироваться, поскольку ее массовый вылов привел к снижению их поголовья.
Итак, вобла и тарань – полупроходные подвиды плотвы обыкновенной, которых ловят в низовьях крупных притоков Каспийского, Азовского и Черного моря, и непосредственно в этих водоемах. Лучшим временем для их ужения является преднерестовый ход в начале весны, но и в остальные периоды на их поимку можно рассчитывать. Главное, правильно выбрать место рыбалки, снасти, приманки и приготовить аппетитную прикормку.
Источники:
http://fishingwiki.ru/%D0%A2%D0%B0%D1%80%D0%B0%D0%BD%D1%8C
http://foodfor.ru/ryba-taran
http://fishelovka.ru/fish/chto-za-ryby-vobla-i-taran
Вяленая рыба: настоящий мужской деликатес. Cушение и вяление рыбы
Богатство вкусу пенным напиткам придает различная закуска — аппетитная соленая и ароматная вяленая рыбка служит идеальным решением для изысканных ценителей гармонии вкусов. Приготовить вяленое лакомство можно практически из любой рыбы.
Приготовить вяленое лакомство можно практически из любой рыбы.
В России исторически сложилось так, что самыми популярными видами рыб для вялки, являются лещ и вобла. Помимо них, есть еще огромное множество рыб, которым следует уделить внимание. При выборе вяленой рыбы, нужно знать все тонкости и нюансы сушения, особенности и секреты процесса вялки, следует располагать необходимой информацией о плюсах и минусах вяленой закуски и прочим — все интересующие моменты описаны ниже.
Особенности процесса вяления — как не прогадать с качеством
Успех процессу вяления задает ряд важных параметров. При вялении в домашних условиях или на производстве, необходимо соблюдать определенные рекомендации:
- Замороженная рыба плохо подходит для процесса вяления. К вялению рекомендуется охлажденное сырье. У замороженной рыбы, при размораживании, частично разрушается мышечное волокно, что приводит к некачественной просолке в дальнейшем. На замороженный продукт уходит гораздо меньше соли, чем на свежий. Вызвано это тем, что при дефростации, из тушки замороженной рыбы выходит немного жидкости.
- Наиболее подходящие условия для вялки рыбы — тень и хорошо проветриваемое место. На солнце рыба выделяет жир, из-за которого, вкус готового продукта сильно портится. Вяленая на солнце рыба имеет желтоватый окрас.
- Крупную рыбу, перед тем, как приступить к сушке, лучше разделать на филе или выпотрошить. Такой подход позволит рыбе качественно провялиться. Некрупных особей можно сушить целиком — рыба хорошо пропитывается жиром, как результат, равномернее просаливается.
Разница между сушеной и вяленой рыбой
Рыбную закуску можно готовить несколькими способами: сушением и вялением. Не зависимо от способа заготовки, в народе такую рыбу называют вяленой. Для сушки и вялки подходит разная рыба, к тому же, данные способы отличаются по заготовке. Различия описаны в таблице:
|
|
Сушеная |
Вяленая |
|
Соль |
Свежая и соленая |
Только соленая |
|
Жирность |
Низкая и средняя жирность |
Жирная |
|
Способ |
Холодный (природный, в прохладную погоду) или горячий (печь) |
Теплый, природный способ (в теплую погоду в тени, в хорошо проветриваемом месте). |
|
Срок |
1,5-2 месяца (холодный способ) или 5-6 часов, быстрый (горячий вариант) |
2-3 недели или немного больше — в зависимости от погоды |
 Рыба предварительно солится.
Рыба предварительно солится.Более мягкой, влажной, прозрачной, получается вяленая рыба. Продолжительная сушка делает рыбу твердой, сухой и соленой. Соответственно, для вяления предпочтительнее выбирать жирную рыбу, для сушки — наоборот. Один и тот же вид рыбы можно приготовить по-разному, здесь все зависит от личных предпочтений человека.
Вяленая рыба: виды и особенности
Для вялки подходит не вся рыба. Хорошо вялятся породы средней жирности. В результате чего, такая рыба получается сухой и соленой, при этом, структура мяса сохраняется. Ниже представлены наиболее популярные виды рыбы для вяления, которые отлично подходят на роль закуски к пенному.
Вяленая вобла (тарань)
Существенного отличия меду воблой и таранью нет — это плотва, которую вылавливают в разных местах. Среди соленых рыбных закусок, самым распространенным продуктом является вобла вяленая. Тарань пользуется популярностью на Кубани и на юге России. Стоимость деликатеса крайне невысока, что делает его доступнее среди обывателей. Мясо такой закуски плотное и сухое, достаточно соленое. Такая рыба отлично подходит к пиву.
Вяленый лещ
Вяленый лещ — речная рыба, традиционная закуска к пиву. Лещ немного крупнее плотвы, не смотря на это, он хорошо подвергается сушке. Вкус у рыбы пикантный.
Вяленая сопа
Рыба семейства карповых — сопа или белоглазка. Особенностью данной рыбы являются ее большие глаза с серебристого цвета радужиной. Мясо в меру жирное, имеет приятный сладковатый вкус.
Вяленый окунь
Окунь, хищная и прожорливая рыбка, имеет белое мясо, которое отличается сравнительно малым числом костей. Полезно мясо окуня тем, что в его составе находится большое количество йода, который оказывает благотворное влияние на человеческий организм. Помимо этого, мясо богато витаминами группы B и содержит большое количество микроэлементов.
Помимо этого, мясо богато витаминами группы B и содержит большое количество микроэлементов.
Вяленая чехонь (сабля)
Чехонь или сабля — рыба семейства карповых, предпочитает стайный образ жизни. Ценится за диетические и вкусовые качества. Мясо данной рыбы имеет приятный аромат и вкус. Вяленая чехонь достаточно жирная.
Вяленая тарашка (густера)
Один из видов плотвы — тарань. Имеет широкое тело и мелкую чешую, крупные зубы и черноватые края плавников.
Вяленая тарань — это традиционно русская закуска к пенному напитку. Вяление и сушка тарани длится от нескольких дней до полумесяца. Длительность приготовления зависит от рецепта, размера рыбы и от желаемого результата. Многим по душе вяленая таранка, некоторые любят сушеную тарань.
Вяленая красноперка
Красноперка входит в семейство карповых, отряд карпообразных. Регион обитания — реки и озера, впадающие в Черное, Аральское, Каспийское, Северное, и Балтийское моря.
Красноперка, по праву, занимает лидирующую позицию, как самая красивая рыбы нашей фауны. Мясо красноперки не слишком жирное, обладает своеобразным вкусом. Мякоть богата витамином Р, хромом и фосфором. Занимательная статистика: На 100 г мякоти данной рыбы приходится 18,5 г белка и 3 г жира.
Блюда из вяленой и сушеной рыбы
Традиционное португальское блюдо из сушеной трески — Бакальяу.
В России вяленую и сушеную рыбу, в том числе и икру, в большинстве своем, употребляют как закуску. В мире вяленая и сушеная рыба выступают в качестве приправы к блюдам или даже, как полноценный ингредиент. Для примера: на азиатской кухне готовят стружку тунца, в Италии — боттаргу (вяленую икру кефали), а в Португалии предпочитают готовить бакальяу( клипфиск, без добавления соли — стокфиск). Последнее добавляется в супы.
Российская кухня тоже тоже богата подобными примерами. На юге страны вяленую рыбу отваривают и употребляют в пищу, как самостоятельное блюдо. Помимо этого, из нее готовят различные супы. На севере страны существует свой традиционный способ заготовки — юккола, схож с португальским стокфисксом. Стокфиск – сушеная треска, без соли.
На севере страны существует свой традиционный способ заготовки — юккола, схож с португальским стокфисксом. Стокфиск – сушеная треска, без соли.
В большинстве своем, привычная россиянину «закусочная» сушеная рыба и икра — это неисчерпаемый кладезь различных кулинарных шедевров. Здесь можно сказать о том, что сушеную рыбу можно использовать в качестве приправы, предварительно мелко нарезав или натерев. Более того, сушеный деликатес можно добавлять в холодные и горячие соуса, в салаты, супы и прочие блюда. Нужно учитывать тот факт, чтобы рыбная нота была уместна в блюде. В качестве нового эксперимента — вяленая рыба является отличным средством для обогащения вкуса того или иного блюда.
Правильное хранение вяленой рыбы
Вяленая рыба, зачастую, может храниться достаточно долго. Для того, чтобы гарантировать сохранность рыбы на протяжении долгого времени, необходимо знать, как правильно хранить сушеную рыбу дома.
Наиболее благоприятные условия для хранения:
- прохладное и сухое место;
- металлическая или стеклянная емкость;
- холодильная камера.
Для каждой породы рыбы обязательно выбирать подходящий способ хранения с целью сохранить вкусовые качества продукта.
Хранить леща лучше всего в деревянной таре, при этом, рыба должна быть обернута в бумагу.
Заморозка лучше всего подходит для воблы и тарани. Перед употреблением, нужно разморозить рыбу, предварительно обернув ее в бумагу, которая сможет впитать излишки влаги.
Корюшка — это та рыба, которую лучше хранить в бумаге, смазав ее маслом. Хранение осуществляется в деревянном ящике.
Важно знать: при комнатной температуре, а также — при повышенной влажности в помещении, вяленая рыба может храниться не более 2х месяцев!
Внимание!
Друзья! Предлагаем купить вяленую рыбку из Астрахани. Вся наша продукция высокого качества, имеет все необходимые документы. Интернет-магазин «Рыба моей мечты» официально зарегистрирован, продукция хранится на складе при необходимом температурном режиме.
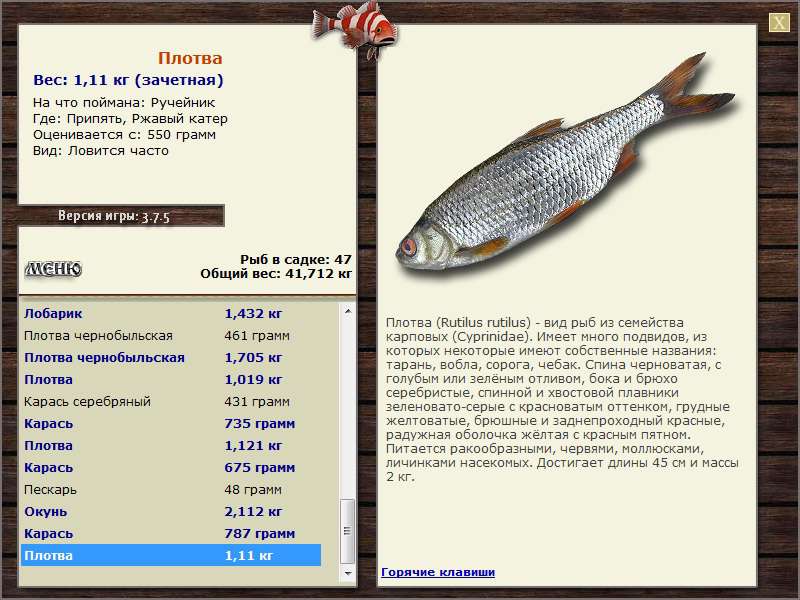
Плотва — Cправочник рыболова.
Плотва, или обыкновенная плотва (Rutilus rutilus) – полупроходная и жилая рыба семейства карповых, широко распространенная в водоемах Европы и Азии. Обыкновенная плотва имеет около 13 подвидов, вот самые известные из них:
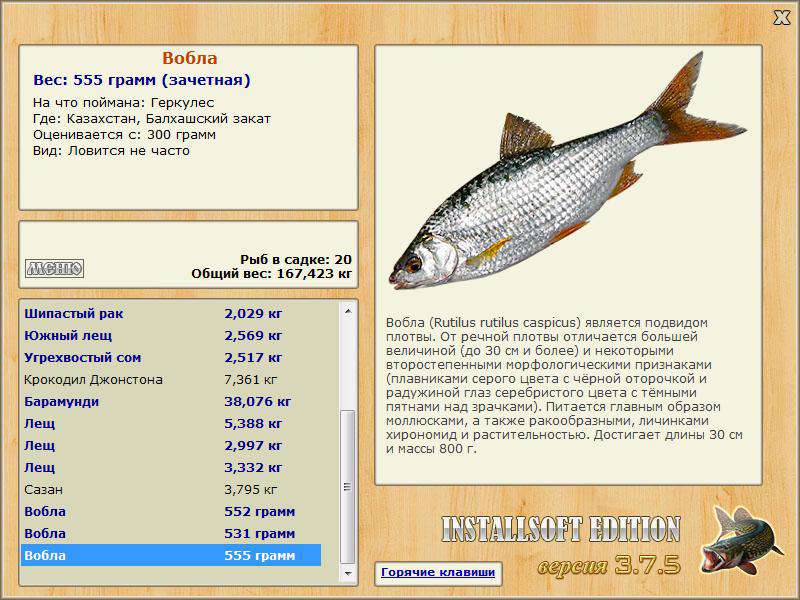
– Тарань, или азовско-черноморская тарань (Rutilus heckelii). Обитает в Черном и Азовском морях, откуда для нереста и зимовки выходит в устья рек.
– Вобла (Rutilus caspicus). Эндемик Каспийского моря.
– Сорога, или сибирская плотва.
Большинство подвидов плотвы, однако, разделены недостаточно четко, поэтому систематика этого вида у многих специалистов вызывает сомнения. Достаточно явными отличиями от обыкновенной плотвы обладает лишь вобла.
Плотва относится к категории промысловых рыб, при этом активнее всего добываются ее полупроходные подвиды – тарань и вобла. Среди жилых форм наиболее ценится сибирская плотва, вылов которой ведется в промышленных масштабах. В продажу рыба поступает в свежем, вяленом и копченом видах.
Внешний вид
Плотва имеет удлиненное, несколько сплюснутое с боков тело, покрытое крупной серебристо-белой чешуей. Спина рыбы при этом имеет темный оттенок, брюхо и бока серебристые, а плавники преимущественно красно-оранжевые. По сравнению с воблой обыкновенная плотва в целом обладает более насыщенным окрасом.
Размеры полупроходных и жилых рыб различаются: если первые вырастают до 51 сантиметра при весе до двух килограммов, то вторые достигают длины не больше 35 сантиметров и массы до 1,3 килограмма.
Места обитания и повадки
Обыкновенная плотва встречается в самых разных водоемах: водохранилищах, реках, прудах, озерах, каналах и не только. Больше всего эта рыба любит участки с водной растительностью и умеренным течением, причем держаться она старается на их границе. Питается плотва различными моллюсками, беспозвоночными и нитчатыми водорослями, а в некоторых случаях поедает мальков и личинок рыб.
Нерест
Половозрелой плотва становится в возрасте 3-5 лет, когда длина ее тела достигает 10-12 сантиметров. Нерест проходит при прогреве воды до 10-12 градусов, что обычно бывает в апреле или в начале мая. Икру самки мечут на растения – от 2,5 до 100 и даже 200 тысяч икринок каждая. Отнерестившись, полупроходные рыбы возвращаются в море, где сразу же приступают к активному питанию.
Через 9-14 дней после икрометания из икры выходят личинки длиной 5-6 миллиметров, которые вскоре начинают питаться мелкими беспозвоночными. Растет плотва довольно медленно, достигая в пятилетнем возрасте массы всего в 80-100 граммов.
описание, класс, питание, места обитания
Плотва — самый распространенный вид после карася, обитающий в различных водоемах. Парочку ее экземпляров можно найти даже в стареющих руслах рек. Не водится она только в закрытых озерах с низким содержанием кислорода.
Что Вы делаете с плотвой (сорожкой)?Poll Options are limited because JavaScript is disabled in your browser.
Это интересно! Образ воблы популярен и используется в кино и мультфильмах: «Большая перемена», «Москва слезам не верит», «Ну погоди!», «12 стульев», «Свадьба в Малиновке» и других.
Описание
Плотва имеет длинное, немного приплюснутое с боков, строение тельца. Спинка черная, с голубоватым или зеленоватым отливом. Серебристая чешуя с достаточно широкими пластинами. Спинной и анальный плавники темные, другие — с оттенком красного. Глаза оранжевые с алым пятном наверху. Пасть оснащена пятью-шестью глоточными зубами.
Во время нереста самец покрывается эпителиальными бугорками («жемчужная сыпь») — брачным нарядом и выглядит шершавым, как напильник.
Жилые особи вырастают до длины 35 сантиметров и веса до 1,3 килограмм. Полупроходные формы крупнее: длина до 51 сантиметра и масса до 2 килограмм. В Чебакуле пойманы экземпляры более значительных размеров.
Сколько живет плотва? Максимальная продолжительность ее жизни достигает 20 лет.
Плотва и сорога
Ученые, занимающиеся исследованием ихтиофауны, выделяют до 13 подвидов плотвички. У каждого из них в разной местности есть отличия в названиях и свои особенности. Рыбакам известны сорога, сорожка, вобла, тарань, бублица, плотица, плотка, красноглазка.
В Сибири и на Урале это узкий чебак с желтыми глазами. В Архангельской, Вологодской, Кировской областях, Ненецком автономном округе — широкая красноглазая сорога. На Байкале и Енисее — сорожка.
То есть, сорога и плотва — это все разновидность одного и того же вида.
Отдельными ихтиологами определен подвид — каспийская вобла (Rutilus caspicus) — морской обитатель Северного Каспия, который в реках только нерестится.
Некорректным будет данное ей в народе имя — «красноперая плотва», в котором смешаны названия двух различных видов: Rutilus rutilus и красноперки Scardinius. Отличия состоят в радужке глаз: плотица — красноглазая, вторая — желтоглазая. Также, у плотвы 10-12 мягких перьев на спинном плавнике, у красноперки — 8-9.
Любопытно! Представитель семейства вырезуб или кутум (на фото ниже) занесен в международную и российскую Красные книги. Ареал его обитания — каспийская водосборная площадь.
Где обитает: морская или речная
Жилая плотва — пресноводная, полупроходная разновидность водится в солоноватых водах.
Растет серебристая рыбка в Евроазиатской части практически во всех озерах, лиманах, водохранилищах, каналах, начиная от Скандинавии и заканчивая Средиземноморьем.
В Центральной части России, в Сибири, на Урале, за исключением Дальнего Востока (ее там нет) она облюбовала:
- Кольский полуостров (Ловозеро, Имандра, реку Поной),
- Карелию (Сямозеро, Выгозеро, Сегозеро и другие),
- реки, впадающие в Северный Ледовитый океан,
- реку Сеяха (полуостров Ямал),
- водоемы Урала,
- бассейны рек Черного, Азовского морей (кроме Крыма),
- системы рек Аральского моря,
- речные бассейны горного Алтая,
- Байкал.
Среда обитания плотвы — пресноводная, с медленным течением. Она держится стайками в заводях, затонах, плесах, старицах и других биотопах с густой растительностью и илистым дном. Дрейфует постоянно на их границах и при опасности скрывается в них.
Важно! В техногенно-загрязненных озерах с талой и дождевой водой из городских территорий вылавливают плотву, у которой строение измененного скелета выглядит аномально: 14 лучей в спинном плавнике, два ряда глоточных зубов, с дополнительными звеньями в позвоночнике. Это значит, что формируется новый морфотип рыбы.
Чем питается плотва
Плотва ест все. Отличается очень разнообразным рационом, но это не значит, что она — хищник. Это травоядный организм.
Молодь и мальки питаются останками беспозвоночных, личинками водных насекомых. В первое лето жизни мальки предпочитают одноклеточные саргассы и дафнии, а осенью ищут мелких водоплавающих.
Сеголетки и зрелая рыба длиной до 15 сантиметров любят полакомиться:
- головастиками,
- комаром-дергуном,
- мелкими навозными червями,
- личинками стрекоз,
- опарышами,
- мальками,
- подлистниками,
- нитчатыми водорослями.
Более крупные экземпляры поглощают пищевых моллюсков: мидий, устриц, гастропод. В глубоких водоемах, где много двухстворчатой речной дрейссены, которых плотва усиленно ест с пяти-шестилетнего возраста, попадаются единицы весом до 1 килограмма.
Первые 3-4 года для плотвы характерен линейный прирост, в последующие периоды жизни — набор массы.
Размножение
Половая зрелость рыбки наступает примерно в три года, отдельные представители являются половозрелыми уже к двухлетию.
Плотва нерестится при раскалывании льда. Обычно это период с конца апреля до середины мая, когда температура воды доходит до 10-12°С, а на мелководье — до 16-17°С. После весеннего очищения водоема ото льда глубина прогревается довольно быстро, и создаются хорошие условия для развития кормовой базы, в особенности планктона, фито- и зообентоса, водной растительности.
Обычно это период с конца апреля до середины мая, когда температура воды доходит до 10-12°С, а на мелководье — до 16-17°С. После весеннего очищения водоема ото льда глубина прогревается довольно быстро, и создаются хорошие условия для развития кормовой базы, в особенности планктона, фито- и зообентоса, водной растительности.
Размножение сопровождается подходом большого количества самцов, которые находятся на территории дольше самок и оплодотворяют икру нескольких из них.
Самки отличаются плодовитостью: одна особь выметывает от трех до ста тысяч икринок сразу.
Плотва является фитофилом: ее продукты метания приклеиваются к плавающей на поверхности старой или свежей растительности (стебли, листья), на корни деревьев, тростника, коряги, камни. Располагаются они меж собой очень тесно, а если находятся на мху, то похожи на виноградные гроздья. Самые крупные скопления содержат в себе более 84 тысяч единиц.
Диаметр икринки составляет 1,5 миллиметра. Она вырастает в личинку за 9 дней — 2 недели. Длина детвы при выклеве достигает 5,2-6,6 миллиметров.
В первые дни жизни личинки еще в покое держатся в местах нереста за стебли и листья при помощи специальных клейковидных выделений из железы на нижней стороне рыла.
Полупроходная плотва крупная и растет более быстрыми темпами. Ее плодовитость достигает 200 тысяч икринок. После нереста взрослые экземпляры возвращаются в море и усиленно откармливается, съедая столько, сколько весят сами.
Занимательно! Перед нерестом рыбка собирается в многочисленные стаи, подходит к кромке водной растительности и постоянно кормится. В этот период ее можно поймать на все, даже на пенопласт.
Естественные враги
В природной среде у этой пугливой рыбицы много врагов.
Весной и в особенности в начале лета значительное количество ее икринок гибнет из-за активного поедания угрями.
Огромная хищная рыба (окунь, щука, судак, прожорливый голавль) постоянно увязывается за косяками тараньки, нападая и в нерестовый период. Она караулит молодь плотвички, заплывающую за планктоном, в подводных зарослях.
Врагом для рыбы являются хищная пернатая чайка с бакланом. Они за один день съедают около 500 грамм. Зимородки предпочитают молодняк и меленькую рыбешку ростом не более 10 сантиметров. Цапли пожирают зрелую плотву размером около 35 сантиметров. Водоплавающие большие поганки — чомги — пасутся в прибрежной зоне для ловли некрупных рыбок.
Особей уничтожает и водоплавающий хищник: норка, выдра, ондатр. Они промышляют ее вдоль побережья. Мелкие экземпляры плотвы съедаются ими тут же в воде, крупногабаритные проглатываются на земле.
Сорожку одолевают и разнообразные недуги, от которых она погибает. Из-за поедания улиток, зараженных паразитическим червем, тарань болеет черно-пятнистым расстройством, когда на теле хворающей рыбы проступают темные бляшки. Для человека инфицированные экземпляры не опасны.
В результате кормления водяными блохами плотва заражается лигулезом. При нем в брюшной полости развиваются ленточные черви, передавливающие внутренние системы. Таранка становится бесплодной и гибнет.
К какому классу и отряду относится
Описание вида — обыкновенная плотва — строится следующим образом:
- надцарство — эукариоты (ядросодержащие клетки),
- царство — животные,
- подцарство — эуметазои (настоящие многоклеточные),
- билатерии (двусторонне-симметричные) — имеется ось тела,
- вторичноротые,
- тип — хордовые,
- подтип — позвоночные,
- инфратип (группа) — челюстноротые,
- клада (группа с общим предком и всеми его потомками) — рыбы,
- надкласс — костные рыбы,
- класс — лучеперые,
- подкласс — новоперые,
- инфракласс — костистые,
- надотряд — костнопузырные (имеется веберов аппарат, который соединяет плавательный пузырь с внутренним ухом),
- серия — остариофизики,
- отряд — карпообразные,
- надсемейство — карпоподобные,
- семейство — карповые (Cyprinidae),
- подсемейство — Leuciscinae (истинные пескари),
- род — плотва,
- вид — плотва.

Международное научное название Rutilus rutilus (Linnaeus, 1758). Имя присвоено шведским биологом Карлом фон Линнеем, сформировавшим биноминальную систему, при которой живым существам даются двойные названия.
Это увлекательно! На свете живет испанская, ширванская, альпийская, итальянская и другие сорожки.
Самая большая
Плотва в мире относится к сорной, и не представляет промышленное значение. В последнее время ловится исключительно для восстановления зоопланктона, благодаря чему ей на смену приходят более ценные сорта рыб. В Европе спросом не пользуется, так как из нее готовят корма для животных, и признана опасной для Балтики.
Сейчас она имеет значение только для любительской рыбалки.
В 2015 году во время ловли английский школьник из Белфаста (Северная Ирландия, на фото ниже) поймал самую большую плотву весом 1,39 кг.
Занимательно! Абсолютный рекордсмен в мире — найденная в Германской Демократической Республике огромная тарань весом 2,58 килограмма. В России в 1931 году поймали самца возрастом 19 лет массой 2,1 килограмма.
Как приготовить
Плотвочку стряпать легко, так как мясо у нее нежнейшее и готовится очень быстро.
Это действительно ценная рыба, в которой в большом количестве содержится кальций, аминокислоты, фосфор.
Единственным недостатком является присутствие в избытке мелких межмышечных косточек. Поэтому, перед жарением на теле рыбы делают острым ножом несколько надрезов. Но из-за них рыбка сохнет и становится не такой сочной. Для придания мягкости ее притушивают с молоком, сметаной, овощами или сохраняют в маринаде из майонеза или другого соуса.
Из тараньки хозяйка может приготовить множество блюд. Суп из нее варить не рекомендуют, так как рыба сильно разваривается, лучше ее жарить, запекать в духовке с соусом и в фольге. Из плотвы получаются очень вкусные котлеты.
Из плотвы получаются очень вкусные котлеты.
Чтобы избавиться от сильного запаха тины, очищенные тушки сороги выдерживают несколько минут в подкисленной воде с лимонной кислотой и уксусом.
Теперь вы знаете все об этой удивительной рыбе!
описание, фото, способы ловли и снасти для плотвы, сезоны
Плотва — подробное описание рыбы и места её обитания. Какой рекордный размер для плотвы. В чем отличия плотвы от воблы и тарани. На что она клюёт?
Содержание:
Родственники
Описание плотвы
Где обитает плотва
Рекордные размеры
На что клюёт
Плотва относится к семейству карповых, и эта рыба является одной из самых распространённых в водоёмах России, причём встречается почти везде, а в реках и озёрах Сибири эта рыба достигает крупных размеров. Только там она называется чебаком. В наших же краях плотву зовут сорожкой, а если рыба крупная, то – сорогой. В отличие от Европы, где плотва также широко распространена, но за рыбу вообще не считается, а идёт в корм скоту и на топливо, в наших же палестинах она очень уважаемая рыба за её красоту, упрямый характер и, если рыба крупная, то и за сопротивление во время вываживания. Кроме того, в российских городах и весях плотву любят за её вкус в вяленом виде. Если она и уступает как закуска к пиву вобле и краснопёрке, то совсем немного. Главное правильно её вялить, но это уже другая история (читайте тут). Кстати, есть ещё одно блюдо, которое при правильном приготовлении оценит самый взыскательный гурман. Это крупная жареная плотва в яйцах. Но рыба должна быть весом не менее трёхсот граммов. Костей, конечно, в ней хватает, как и в леще, но наши люди умеют есть речную рыбу и ценят её.
Родственники плотвы
Близкими родственницами плотве являются вобла и тарань. Отличаются эти рыбы более узким телом и жёлтыми, а не красными глазами, как у плотвы-сорожки. Плотва, конечно, красивее и уступает только краснопёрке, которая также относится к близко родственному плотве виду. Чебак тоже является плотвой, но по виду больше похож на воблу и тарань. Вобла, тарань и чебак обычно более крупные и килограммовый вес этих рыб не является предельным и редким, что для плотвы можно назвать трофейным весом. Кроме того, указанные выше рыбы более жирные. Это связано с другим видом питания. Если плотва летом переходит на растительную пищу, куда входит и зелень (шелковник), которой бывает просто переполнена эта рыба и нередко из-за этого горчит, то вобла, тарань и чебак не прочь закусить и моллюсками, мелкими ракообразными, крупными насекомыми. Более калорийный вид питания делает мясо этих рыб также более калорийным и жирным.
Описание плотвы и её отличие от воблы и тарани
Отличие плотвы от воблы и тарани состоит также в том, что сорога рыба оседлая. А вобла и тарань относятся к полупроходным видам рыб, которые обитают и на опреснённых участках морей, то есть в устьях рек, впадающих в Каспийское и Аральское моря.
К основным отличиям плотвы от родственных видов рыб относятся незазубренные глоточные зубы в один ряд, где на каждую сторону приходится по 5-6 зубов. Кроме того, чешуя плотвы более крупная. Спинка тёмная с зелёным оттенком. Плавники поверху серо-зелёные с красноватым оттенком. По нижней части – красные. Чешуя серебристая, иногда с синевой и нередко с золотистым оттенком, если плотва является гибридом с краснопёркой. Радужка глаз обычно золотисто-жёлтая с красным пятнышком. Словом, красивая рыбка. От краснопёрки плотву отличает ещё то, что у неё мягких перьев спинного плавника бывает от десяти до двенадцати. А у краснопёрки – восемь-девять перьев.
Где обитает плотва
Плотва-сорога предпочитает тихие укромные места, заросшие растительностью, с умеренным течением. Любит, когда над водой находятся ветви деревьев, с которых падают насекомые. Также держится и в коряжистых местах, где топляки и торчащие из воды сучья обрастают зеленью-шелковником – летним кормом плотвы. Плотва – рыба преимущественно стайная и если уже начинает клевать, то берёт иногда одна за другой.
Весной плотва поднимается на нерест в верховья малых рек, где нерестится на мелководьях, заросших растительностью. Этот весенний ход плотвы считается наилучшим временем для её ловли, когда обычно используются донные снасти, в том числе фидер и закидушки (о ловле плотвы весной). В это время обычно попадаются самые крупные экземпляры этой рыбы.
Рекордные размеры плотвы
Рекордный для плотвы размер – 50 см при весе в 3 кг. Это её предельные параметры при максимальной продолжительности жизни в двадцать один год. В наше время самые крупные рыбины были пойманы в Германии – 2,58 кг, а в Финляндии – 2,550 кг.
В летнее время наиболее распространёнными снастями для ловли плотвы-сороги считаются поплавочные удочки разных видов. Для ловли на малых реках у травы используется маховая удочка. Для дальнего заброса применяются болонская и матчевая удочки.
На что клюёт плотва
Плотва клюёт на червя, опарыша, мотыля, манку, хлеб, перловку, кукурузу, зелень.
Рыбное – Плотва
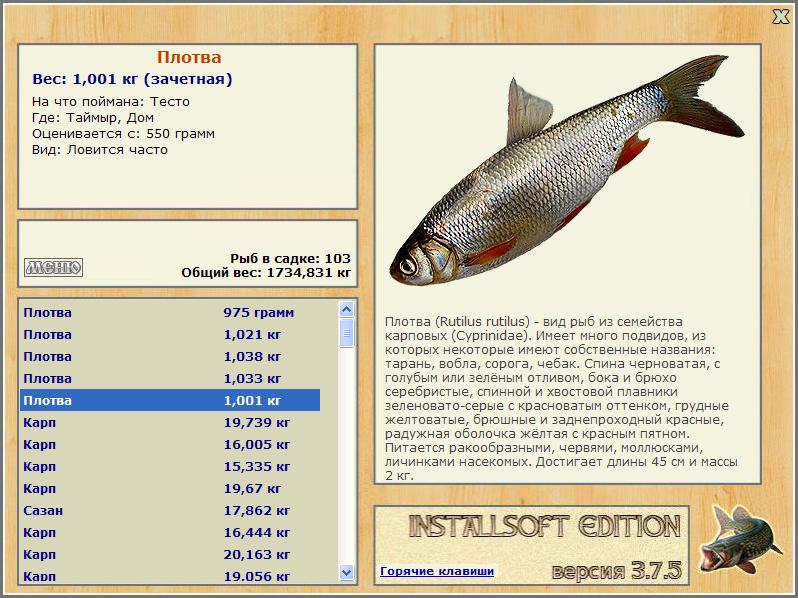
Плотва? обыкнове?нная (плотва, тара?нь, во?бла, соро?га) (лат. Rutilus rutilus) — вид рыб из семейства карповых (Cyprinidae). Имеет много подвидов, из которых некоторые имеют собственные названия: тарань, вобла, сорога.
В Сибири, как минимум в Западной её части, и на Урале общеупотребительное название плотвы — чебак. Чебак с желтыми глазами и узкий, Сорога – с красными глазами и широкая. В Архангельской области общеупотребительное название для обыкновенной плотвы — «сорога».
Плотва встречается по всей Европе к востоку от Южной Англии и Пиренеев и к северу от Альп; в реках и озёрах Сибири, в бассейнах Каспийского и Аральского морей.
От ближайших к ней видов плотва отличается незазубренными и расположенными с каждой стороны в один ряд глоточными зубами (по 5—6 с каждой стороны), относительно крупной чешуей (40—45 чешуй в боковой линии), пастью на конце морды и положением начала спинного плавника над основаниями брюшных. Спина черноватая, с голубым или зелёным отливом, бока и брюхо серебристые, спинной и хвостовой плавники зеленовато-серые с красноватым оттенком, грудные желтоватые, брюшные и заднепроходный красные, радужная оболочка жёлтая с красным пятном. Попадаются также экземпляры с глазами и плавниками жёлтого цвета, с золотистой чешуей, с красноватым оттенком на боках и спине.
Наиболее простой способ отличия от краснопёрки — цвет глаз: у краснопёрки — глаза оранжевые, с красным пятном вверху, тогда как у плотвы — кроваво-красный. Ещё одно отличие — количество мягких перьев на спинном плавнике: у краснопёрки их 8—9, тогда как у плотвы 10—12. Иногда встречаются гибридные формы этих рыб, обладающие признаками обоих видов.
Держится обычно стаями в местах со слабым течением под защитой коряг, свисающих деревьев или водной растительности. При этом в стае средних и мелких рыб могут быть и единичные крупные экз. Мелкая и средняя рыба не пуглива.
С наступлением холодов, как и подавляющее большинство мирных рыб, она собирается в крупные стаи и перекочевывает в глубоководные заводи с замедленным течением. Здесь держится в течение всей зимы, в оттепели отдельные ее особи могут подходить к берегам в поисках пищи. После вскрытия водоемов и с подъемом паводковых вод плотва покидает зимние убежища и выходит в залитую водой пойму и в пойменные водоемы. При прогреве воды до 8° и выше плотва начинает нереститься. Наиболее активно происходит икрометание ее при температуре 10-12°. Во время нереста она образует стаи, которые подходят в тихие, сильно заросшие прибрежные участки водоемов. Процесс икрометания протекает бурно. В тихую погоду, на предвечерней заре и особенно с восходом солнца постоянно слышны всплески воды в результате нерестовых игр производителей, некоторые из них выскакивают над поверхностью воды. Самцы перед нерестом покрываются жемчужной сыпью, придающей шероховатость их телу. Вскоре после нереста этот брачный наряд исчезает. Икру плотва откладывает на прошлогоднюю растительность (стебли, листья), на плавающие ее остатки, а также на корни ив, тростника и других растений. Течение воды на местах нереста отсутствует или же его скорость не превышает 0,2- 0,3 м/сек.
Процесс икрометания протекает бурно. В тихую погоду, на предвечерней заре и особенно с восходом солнца постоянно слышны всплески воды в результате нерестовых игр производителей, некоторые из них выскакивают над поверхностью воды. Самцы перед нерестом покрываются жемчужной сыпью, придающей шероховатость их телу. Вскоре после нереста этот брачный наряд исчезает. Икру плотва откладывает на прошлогоднюю растительность (стебли, листья), на плавающие ее остатки, а также на корни ив, тростника и других растений. Течение воды на местах нереста отсутствует или же его скорость не превышает 0,2- 0,3 м/сек.
Как ловить
Снасть для ловли плотва ничего необычного не содержит. Спиннинговое удилище для осуществления заброски, леска или шнур, на конце лески кормушка с кормом, отвод и поводок с крючками № 4 – № 6 по отечественной классификации. Велосипед изобретать не надо. Подробнее коснемся приманок и техники ловли. Именно от них часто зависит результат рыбалки. Многие рыбаки идеально подбирают снасти, прикорм, приманки, но при этом забывают об элементарных правилах самой ловли. Начнем, пожалуй, с выбора приманок. В качестве приманок хорошо себя зарекомендовали красные черви и мастирка. Перловка, пусть даже окрашеная в желтый цвет, пусть даже сдобренная ароматизатором является третьесортной приманкой. Они должны хорошо держаться на крючке, прежде всего во время заброски. А потом уже и во время поклевок плотвы. Мастирка, как и любая другая растительная приманка, по этому критерию уступает красным червям. Очень часто растительные приманки просто сбиваются при ударе крючка о воду. Поэтому, если с заброской у вас пока туговато, лучше доверять червям. Но с другой стороны мастирка плотве нравится больше. Исключения, конечно, тоже есть, но с мастиркой результат все же лучше. А теперь остановимся на технике ловли. Прежде всего разберемся в том, как же наша приманка попадает в пасть к плотве. Стайки плотвы снуют в толще воды и хватают любого червячка, личинку и другую мимо проплывают живность. Но привлечь целую стайку одним шариком мастирки или одним червячком просто не получится. Это не шаловливая верховодка и не жирующий окунек, хватающие все, что находится в поле зрения. У плотвы, как в принципе и у любой рыбы, есть в воде уже знакомые объекты, которые она схватит. К ним относятся живущие в воде личинки комаров и другие червячки. Все остальное плотва, при наличии привычного ей корма, проигнорирует. Этот привычный корм часто лежит на дне, и в сплетня есть возможность хорошенько его рассмотреть и распробовалы. Плюс, если корм найден, плотва в этом месте и останется. И здесь свою роль играет Кормак. При падении на дно корм вымывается, создавая муть. Эта муть плотва очень интересует, и привлекает рыбу к приманке. А потом начинается дегустирование – примет червячка в рот, вылюнет, за кончик потянет. И через некоторое время, наконец-то его заглотит. И далеко не факт, что приманка к тому моменту еще останется на крючке. Особенно, если это мастирка. В отличие от леща, плотва не втягивает весь корм, как насос, а пытается хорошенько все распробовать.
Но привлечь целую стайку одним шариком мастирки или одним червячком просто не получится. Это не шаловливая верховодка и не жирующий окунек, хватающие все, что находится в поле зрения. У плотвы, как в принципе и у любой рыбы, есть в воде уже знакомые объекты, которые она схватит. К ним относятся живущие в воде личинки комаров и другие червячки. Все остальное плотва, при наличии привычного ей корма, проигнорирует. Этот привычный корм часто лежит на дне, и в сплетня есть возможность хорошенько его рассмотреть и распробовалы. Плюс, если корм найден, плотва в этом месте и останется. И здесь свою роль играет Кормак. При падении на дно корм вымывается, создавая муть. Эта муть плотва очень интересует, и привлекает рыбу к приманке. А потом начинается дегустирование – примет червячка в рот, вылюнет, за кончик потянет. И через некоторое время, наконец-то его заглотит. И далеко не факт, что приманка к тому моменту еще останется на крючке. Особенно, если это мастирка. В отличие от леща, плотва не втягивает весь корм, как насос, а пытается хорошенько все распробовать.
Как же сделать так, чтобы плотву можно было чаще засекать? После долгих распробований мастирки или червячка приманка может просто не удержаться на крючке, и около половины подсечек окажутся холостыми. Чтобы этого избежать, нужно правильно цеплять приманку. Мастирку (или другую растительную приманку) насаживается на крючок так, чтобы острие крючка упирается в поверхность «мастирного» шарики. Мастирку насаживается НЕ шариком, а в виде конусообразного мешочка. Т.е. нижнюю часть делаем толще, верхнюю тоньше, а на конце, в верхней точке мешочка, совсем острый конец. Вот в этот острый наконечник и должно вплотную упираться острие крючка. Плюс такой «мешочек» гораздо лучше держится на крючке. Червей насаживать нужно подобно чулкам – просто надевать их на крючок. И чтобы острие также упирался в тело червя, но ни в коем случае не было открыто. Это же относится и к мастирке. Теперь рассмотрим поклевку.
Когда плотва заметит нашу приманку и подплывет, начнется долгий процесс «дегустирования». Минута этого процесса – и из-за небрежности в насаживании поклевка может прекратиться. Плотва, наверное, напоролись на острие, или хуже того, кусочек шарика с мастирки отломился, обнажив острие. Тогда плотва бросает приманку. Но если плотва напорется на острие в нашем случае, она засечется. Здесь уже нас должны выручить хорошие колокольчики, чтобы вовремя заметить поклевку, и зрение, чтобы вовремя подсечь рыбу. Уверенное натяжение лески – следует подсечка. Следуя этим несложным правилам, количество пойманной рыбы заметно возрастет. Мелкую плотву, до 300г весом, сейчас можно вполне неплохо ловить возле берега на поплавочную удочку на те же приманки (разумеется, без Кормак). Мелкие плотвички не так капризны, как их старшие собратья, и очень часто рыбалка проходит без скуки. Не пропустите весенний ход плотвы.
Минута этого процесса – и из-за небрежности в насаживании поклевка может прекратиться. Плотва, наверное, напоролись на острие, или хуже того, кусочек шарика с мастирки отломился, обнажив острие. Тогда плотва бросает приманку. Но если плотва напорется на острие в нашем случае, она засечется. Здесь уже нас должны выручить хорошие колокольчики, чтобы вовремя заметить поклевку, и зрение, чтобы вовремя подсечь рыбу. Уверенное натяжение лески – следует подсечка. Следуя этим несложным правилам, количество пойманной рыбы заметно возрастет. Мелкую плотву, до 300г весом, сейчас можно вполне неплохо ловить возле берега на поплавочную удочку на те же приманки (разумеется, без Кормак). Мелкие плотвички не так капризны, как их старшие собратья, и очень часто рыбалка проходит без скуки. Не пропустите весенний ход плотвы.
5 распространенных видов тараканов, которые могут оказаться в вашем доме
ПоравутеГетти Изображений
Если вы никогда не видели таракана у себя дома, считайте, что вам повезло. И если да, то вы знаете, сколько боли они могут доставить – как найти, так и избавиться от них. Но, как и другие насекомые, существует несколько различных типов тараканов, которые могут проникнуть в ваш дом, и в зависимости от того, какой из них вы обнаружите, на самом деле определит, как от них лучше всего избавиться.
В целом тараканы делятся на две основные группы: домашние и перидоместные тараканы. Домашние тараканы были с нами со времен пещерного человека и со временем эволюционировали, чтобы жить строго в среде человека, – говорит Дини Миллер, доктор философии, профессор городской энтомологии в Технологическом институте Вирджинии и специалист по борьбе с городскими вредителями в штате Вирджиния. «У нас нет животных, которые бегают вокруг этих парней», – говорит она.
Вторая группа – перидоместные, в которую входят тараканы, которые привыкли находиться рядом с людьми, но не обязательно должны жить в наших жилищах, – говорит Миллер. «Они будут приходить на помощь», – говорит Миллер. «Допустим, на улице слишком жарко или слишком сухо. Эти тараканы в конечном итоге попадут в здания только потому, что там, где они были раньше, сейчас не работает ».
«Они будут приходить на помощь», – говорит Миллер. «Допустим, на улице слишком жарко или слишком сухо. Эти тараканы в конечном итоге попадут в здания только потому, что там, где они были раньше, сейчас не работает ».
Но внутри этих групп вы найдете еще больше видов тараканов, от некоторых из которых избавиться значительно труднее, чем от других. Вот как отличить их друг от друга и что вам нужно знать, чтобы официально выселить этих нежелательных соседей по комнате.
Реклама – продолжить чтение ниже
Немецкий таракан
Как они выглядят: Немецкий таракан – это небольшая, светло-коричневая плотва в форме торпеды, длина которой обычно не превышает says дюйма, – говорит Коби Шаль, доктор философии, возглавляющий лабораторию Schal Lab в Северной Каролине. Государственный университет, специализирующийся на изучении тараканов. «Немецкий таракан очень отличается тем, что я называю гоночными полосами», – говорит Шаль.Взрослые немецкие тараканы имеют две полосы на переднеспинке – пластине за головой – которые вы не найдете у других видов тараканов.
Где их найти: Немецкий таракан – наиболее распространенный вид плотвы, который живет в домах и размножается в них, говорит Шаль. Но, как правило, немецкий таракан чаще всего заражает среду, похожую на городскую. «Этот таракан действительно может размножаться в таких условиях, особенно в условиях с низким уровнем дохода, где у людей нет ресурсов, чтобы тратить много денег на борьбу с тараканами», – говорит Шал.
«Когда мы говорим о государственном жилье и людях, живущих с тысячами домашних тараканов, мы всегда говорим по-немецки», – добавляет Миллер. Немецкие тараканы очень зависят от воды, поэтому, если они есть у вас дома, вы, скорее всего, найдете их на кухне или в ванной.
Как от них избавиться : К сожалению, немецкие тараканы выработали устойчивость ко многим инсектицидам. Это означает, что вам нужно проявить немного больше активности, чтобы избавиться от них. Миллер рекомендует делать «тако с приманкой» – в котором вы помещаете линию приманки для тараканов в центре сложенного 2-дюймового квадрата вощеной бумаги – и меняйте тип приманки, которую вы используете в этом тако каждые три месяца, чтобы он всегда имеет другой тип действующего вещества.Таким образом, вы нацелитесь на всех немецких тараканов, которые могут быть в вашем доме, даже на тех, которые могут иметь устойчивость к определенным типам инсектицидов.
Миллер рекомендует делать «тако с приманкой» – в котором вы помещаете линию приманки для тараканов в центре сложенного 2-дюймового квадрата вощеной бумаги – и меняйте тип приманки, которую вы используете в этом тако каждые три месяца, чтобы он всегда имеет другой тип действующего вещества.Таким образом, вы нацелитесь на всех немецких тараканов, которые могут быть в вашем доме, даже на тех, которые могут иметь устойчивость к определенным типам инсектицидов.
МАГАЗИН COCKROACH BAIT
Коричнево-полосатый таракан
Как они выглядят: Таракан с коричневой полосой – один из самых маленьких тараканов, наряду с немецкой плотвой. «Особенно с коричневыми полосами, если они незрелые, вы можете даже не догадываться, что это таракан», – говорит Скотт О’Нил, доктор философии.D., исследователь городской энтомологии из Университета Небраски. По его словам, он также обычно имеет красновато-коричневый цвет.
Где вы их найдете: В отличие от низких трещин и щелей в ванных комнатах или на кухнях, где обычно встречаются немецкие тараканы, тараканы с коричневыми полосами обычно находятся выше в комнатах, поскольку у них нет такая же потребность в воде, – говорит Миллер.
Одна хорошая вещь, на которую стоит обратить внимание? «Несмотря на то, что они живут с нами и никому не нравятся, на самом деле они не такие вредители, как немецкие тараканы, которые ползают по нам, ползают по нашей еде и тому подобное», – говорит Миллер.
Как от них избавиться: Коричневые тараканы ни к чему не устойчивы, поэтому их убьет любой тип наживки, говорит Миллер. Это означает, что вы, вероятно, можете просто делать одно или два применения с одной и той же приманкой, а не вращать приманки каждые несколько месяцев.
Американский таракан
Как они выглядят: Американский таракан – самый крупный таракан в Северной Америке, его рост составляет около 2 дюймов во взрослом возрасте, – говорит Шаль. Он также имеет блестящий и красноватый цвет, и его часто называют «жук пальметто».
Он также имеет блестящий и красноватый цвет, и его часто называют «жук пальметто».
Где их найти: Американские тараканы чаще всего встречаются в городских квартирах на нижних этажах, говорит Шаль, потому что этот тип тараканов обитает в основном в канализационных системах.
«Во многих городах я думаю о них как о плавающих на циновке из тараканов, потому что под нами огромная канализационная сеть, и эта канализационная сеть обычно занята миллионами этих тараканов», – говорит Шаль.Американские тараканы, особенно летом, часто выходят из канализационной системы, спасаясь от жары и ища пищу.
Как от них избавиться: Просто нанесите несколько капель наживки в том месте, где вы находите американских тараканов в своем доме, – говорит Шал. Скорее всего, это будет в кухонных ящиках, за холодильником или под раковиной.
Дымный коричневый таракан
Как они выглядят: Дымчато-коричневые тараканы по размеру схожи с американскими тараканами – достигают 2 дюймов, но имеют почти черный цвет, – говорит Шаль.Хотя они к тому же блестящие, у них нет такого блеска, как у американского таракана.
Где их найти: Если вы живете в пригороде и обнаружили у себя дома таракан, скорее всего, виноват дымчато-коричневый. Хотя этот вид тараканов не будет активно пытаться проникнуть в ваш дом, в зимние месяцы его часто привозят с дровами. «Они обычно проводят зиму в бревнах», – говорит Шаль. «И когда мы приносим вещи в дом, мы, как правило, приносим их.”
Как от них избавиться: Этот тип тараканов обычно не заражается вашим домом, потому что он вообще не хочет находиться в вашем доме”, – говорит Шаль. Тем не менее, попробуйте убить таракана, которого вы видите, не используя сначала инсектициды (подумайте: обувь или молоток). Если после этого вы все еще видите тараканов, пора вывести наживку.
Восточный таракан
Как они выглядят: Восточный таракан, которого обычно называют «водяным жуком», является крупным и темно-коричневым цветом. Одним из отличительных факторов восточных тараканов является то, что, в отличие от других видов тараканов, у самок нет крыльев, а у самцов только очень короткие, говорит Миллер, поэтому вы вряд ли увидите летающих тараканов этого типа.
Одним из отличительных факторов восточных тараканов является то, что, в отличие от других видов тараканов, у самок нет крыльев, а у самцов только очень короткие, говорит Миллер, поэтому вы вряд ли увидите летающих тараканов этого типа.
Где их найти: Восточные тараканы обычно обитают в более прохладных и влажных местах, говорит Миллер. В одном месте, в котором вы могли бы встретить восточную плотву? Ваш подвал. Это особенно актуально после периодов проливных дождей, так как во время этих ливней восточные тараканы пробираются внутрь.
Как от них избавиться: Как и от большинства тараканов, лучший способ избавиться от восточных тараканов – это положить наживку в том месте, где вы их видели. За исключением того, что в отличие от других типов, это, вероятно, будет означать, что приманку нужно положить в подвал, а не в такие места, как кухня или ванная.
–
Понравилось то, что вы только что прочитали? Вам понравится наш журнал! Зайдите сюда , чтобы подписаться. Не пропустите ничего, загрузив Apple News здесь и выполнив функцию «Профилактика».О, , и мы тоже в Instagram.
Этот контент создается и поддерживается третьей стороной и импортируется на эту страницу, чтобы помочь пользователям указать свои адреса электронной почты. Вы можете найти больше информации об этом и подобном контенте на сайте piano.io.
Реклама – продолжить чтение ниже
КАК МЫ ЖИВЕМ СЕЙЧАС: 7-25-04: ЯВЛЕНИЕ; Неудачная плотва
Точных данных о популяциях плотвы мало, но появляется все больше свидетельств того, что широкое распространение наживки в 90-х годах сказалось.В Майами, традиционно теплом, влажном оплоте тараканов, управление государственного жилищного строительства увидело, что его заказы на работу с тараканами упали с тысяч в год до нескольких сотен в год сегодня. Десятилетний обзор 55 федеральных зданий показал, что количество жалоб на тараканов в период с 1988 по 1999 год снизилось более чем на 93 процента. И хотя тараканы долгое время приносили наибольший доход коммерческим истребителям, по данным отраслевого журнала Pest Control, они опустились на 3-е место после муравьев и термитов.
Десятилетний обзор 55 федеральных зданий показал, что количество жалоб на тараканов в период с 1988 по 1999 год снизилось более чем на 93 процента. И хотя тараканы долгое время приносили наибольший доход коммерческим истребителям, по данным отраслевого журнала Pest Control, они опустились на 3-е место после муравьев и термитов.
Такая же картина сложилась и на потребительском рынке. Продажи готовой наживки резко выросли в 90-х годах до 80 миллионов долларов в год. Потом выпало дно. Рынок всех продуктов для борьбы с тараканами – не только приманок, но и более распространенных аэрозолей от тараканов – начал сокращаться в 1996 году, согласно данным розничной торговли, приведенным компанией Clorox, которая теперь владеет Combat. «Похоже, что они могли быть настолько успешными, что иссушили рынок», – говорит Кен Харрис, основатель Cannondale Associates, консалтинговой фирмы по маркетингу.
К концу 2000 года служба борьбы с вредителями обнаружила, что ведет колонку под названием «Неужели приманки для тараканов слишком эффективны?» Потребительский рынок средств борьбы с тараканами в США продолжает сокращаться на 3-5 процентов в год, говорит Дерек Гордон, вице-президент. президент по маркетингу в Clorox. «И если нам действительно удастся полностью вывести себя из бизнеса, честно говоря, мы почувствуем, что оказали миру услугу».
Однако передышка может быть недолгой. Некоторые операторы по борьбе с вредителями утверждают, что инвазивные виды муравьев бросились заполнять домашнюю нишу, оставленную тараканами.Другие говорят, что, когда операторы превратили остаточные спреи в наживку, они сделали нас все более уязвимыми для постельных клопов, которые в последние несколько лет вернулись, чтобы сосать кровь как в блошиных, так и в пятизвездочных отелях.
Более того, немецкая плотва может снова подняться. На Национальной конференции по городской энтомологии в Фениксе в мае этого года симпозиум-блокбастер имел готовое название: «Воскрешение немецкого таракана». Энтомологи читали статьи и показывали видеоролики, свидетельствующие о том, что тараканы все еще не выработали устойчивость к гидраметилнону. даже после 20 лет интенсивного использования некоторые из них отказались от приманок, содержащих определенные сахара или добавки.
Энтомологи читали статьи и показывали видеоролики, свидетельствующие о том, что тараканы все еще не выработали устойчивость к гидраметилнону. даже после 20 лет интенсивного использования некоторые из них отказались от приманок, содержащих определенные сахара или добавки.
Полученные данные не предвещают возвращения Скенектади. Большинство потребительских приманок по-прежнему работают; и Bayer, химический и фармацевтический гигант, представил формулу, которая удовлетворяет привередливые вкусы новых тараканов. Но у городских энтомологов явно есть веские причины вернуться к тараканам.
«Я до смерти взволнован, что они вернулись», – сказала Дини Миллер на мероприятии. Ученые-тараканы в зале аплодировали.
Границы | Тараканы проявляют индивидуальность в обучении и памяти во время классического и оперантного кондиционирования
Введение
Поведенческий синдром определяет последовательное поведение человека, которое коррелирует во времени и контекстах.Личность животного (Gosling and Vazire, 2002) выражается в долгосрочных различиях между людьми по множеству поведенческих черт, таких как смелость-застенчивость, избегание исследования, уровень активности, общительность или агрессия (Sih et al., 2004a, b. ; Dingemanse, Wolf, 2010). В то время как устойчивые поведенческие черты были тщательно изучены у позвоночных, литературы по индивидуальности и личности у беспозвоночных по-прежнему мало (для обзора см. Kralj-Fišer and Schuett, 2014). Небольшое количество доступных данных о личности беспозвоночных может быть частично связано с традиционным представлением о том, что беспозвоночные проявляют стереотипное поведение стимула-реакции с небольшими индивидуальными различиями (например,г., Brembs, 2013). Исследования беспозвоночных в основном проводились в контексте коллективного поведения в социальных контекстах и в основном изучали, как отдельные личности влияют на поведение колонии (например, у тараканов: Planas-Sitjà et al., 2018; Planas-Sitjà and Deneubourg, 2018, муравьи: Pinter-Wollman, 2012, пауки: Grinsted et al. , 2013; Wright et al., 2014, гороховая тля: Schuett et al., 2011; сверчки: Rose et al., 2017).
, 2013; Wright et al., 2014, гороховая тля: Schuett et al., 2011; сверчки: Rose et al., 2017).
На уровне познания животных различия в способностях индивидуума могут отражать различия в когнитивных способностях независимо от характера животного.Однако индивидуальные когнитивные стили также могут влиять на личность (Carere and Locurto, 2011). Индивидуальность интенсивно изучалась в обучении и памяти. Изучение и запоминание значимости сигналов окружающей среды имеет большое значение для выживания практически всех животных. Как было показано для обоих позвоночных, особи одного вида могут существенно различаться по своим способностям к обучению (например, Gosling, 2001; Gallistel et al., 2004; Groothuis and Carere, 2005; Kolata et al., 2005; Schuett and Dall, 2009). ; Котршал, Таборский, 2010; Дэвид и др., 2011) и беспозвоночных (см. Обзор Dukas, 2008).
Среди насекомых исследования были сосредоточены на шмелях и медоносных пчелах. Шмелей изучали для решения множества задач (Chittka et al., 2003). Например, отдельные шмели, которые изучают только один параметр цветка (запах или цвет), были более эффективны по нескольким причинам, чем те, которые выучили два: они делали меньше ошибок, имели более короткое время обработки цветов, быстрее исправляли ошибки и переходы между цветами были изначально более быстрые (Chittka, Thomson, 1997).Кроме того, было показано, что отдельные шмели постоянно различаются по своей способности научиться различать стимулы по визуальной и обонятельной модальности (Muller and Chittka, 2012). Систематический анализ классических экспериментов по кондиционированию медоносных пчел показал, что среднее по группе обучающее поведение неадекватно отражает поведение отдельных животных. Этот результат соответствовал большому количеству наборов данных, включая обонятельное и тактильное кондиционирование, собранное у более чем 3000 пчел, полученных во время абсолютного и дифференциального классического кондиционирования (Pamir et al., 2011, 2014). Постепенно увеличивающиеся кривые обучения отражали артефакт группового усреднения, а поведенческие характеристики людей характеризовались резким и часто ступенчатым увеличением уровня реакции (Pamir et al., 2011), что напрямую соответствует наблюдениям у позвоночных ( Gallistel et al., 2004), но противоречит более ранним данным, полученным на плодовой мушке ( Drosophila melanogaster ), у которых было описано, что среднее по группе поведение представляет собой вероятностное выражение поведения у отдельных особей (Quinn et al., 1974).
Постепенно увеличивающиеся кривые обучения отражали артефакт группового усреднения, а поведенческие характеристики людей характеризовались резким и часто ступенчатым увеличением уровня реакции (Pamir et al., 2011), что напрямую соответствует наблюдениям у позвоночных ( Gallistel et al., 2004), но противоречит более ранним данным, полученным на плодовой мушке ( Drosophila melanogaster ), у которых было описано, что среднее по группе поведение представляет собой вероятностное выражение поведения у отдельных особей (Quinn et al., 1974).
В настоящей работе мы спрашивали, проявляют ли тараканы индивидуальность в своих способах обучения как в классических, так и в оперантных задачах кондиционирования. Исследования поведенческого обучения, в которых использовались обонятельные и визуальные сигналы, показали, что тараканов можно тестировать на выполнение классических задач кондиционирования, пока животные находятся в иммобилизации (Watanabe et al., 2003; Kwon et al., 2004; Lent and Kwon, 2004; Watanabe and Mizunami, 2006). или могут свободно передвигаться (Watanabe et al., 2003; Sato et al., 2006; Hosono et al., 2016). В некоторых экспериментах после классического обонятельного кондиционирования тесты памяти проводились на открытой арене, где тараканы могли свободно выбирать подход к различным запахам (Watanabe et al., 2003; Sato et al., 2006). Открытые арены и Т-образные лабиринты успешно используются для оперирования тараканами. Балдеррама (1980) впервые продемонстрировал, что тараканов можно обучить ассоциировать разные запахи с сахаром или солевым раствором на открытой арене. Mizunami et al. (1998) изучали память мест с помощью пространственного теплового лабиринта с визуальными подсказками и без них.Более поздние исследования Мизунами и его коллег (Сакура и Мизунами, 2001; Сакура и др., 2002) подтвердили и расширили функциональную обусловленность тараканов на открытой арене. Barraco et al. (1981) успешно обучили тараканов задаче пространственного различения, используя электрический шок, чтобы наказать либо левый, либо правый поворот в Т-образном лабиринте. Используя стимулы различных модальностей, мы показываем в настоящем исследовании, что тараканы демонстрируют индивидуальность в своей способности учиться и запоминать стимулы, используя как классические, так и оперантные задачи обуславливания.
Используя стимулы различных модальностей, мы показываем в настоящем исследовании, что тараканы демонстрируют индивидуальность в своей способности учиться и запоминать стимулы, используя как классические, так и оперантные задачи обуславливания.
Материалы и методы
Насекомые
Все эксперименты проводились с взрослым самцом Periplaneta americana . Животных содержали в условиях обратного цикла свет-темнота (12 ч: 12 ч) при 26 ° C в лабораторных колониях в наших центрах разведения в Кельнском университете. Тараканам разрешалось пить воду и кормить овсом ad libitum . Однако воду удалили за 4 дня до тренировки для повышения мотивации. Все эксперименты проводились в активной фазе (скотофазе) животных.
Экспериментальные установки
Для классического кондиционирования тараканов запрягали в изготовленные на заказ фиксирующие цилиндры (рис. 1А) после анестезии при 4 ° C. После фиксации выступала только голова животного, что позволяло свободно перемещать антенны и ротовой аппарат. После привыкания в экспериментальной комнате в течение 16 часов тараканов помещали перед пластиковым шприцем на 10 мл, установленным в держателе, и вытяжной системой за удалением насыщенного запахом воздуха (рис. 1А). Стимулы разбавляли минеральным маслом (Acros Organics ™, Geel, Бельгия), и концентрацию запаха доводили так, чтобы давление паров запаха соответствовало наименьшему значению (транс-коричный альдегид).Разведения были следующими (в% об. / Об.): Изоамилацетат (99+%, чистый, Acros Organics ™, Geel, Бельгия): 26,27%, масляная кислота (> 99%, Aldrich, Дармштадт, Германия): 2,56%, транс-коричный альдегид (≥ 98%, Merck, Дармштадт, Германия) неразбавленный. Десять микролитров каждого запаха вводили на кусок фильтровальной бумаги, вставленный в пластиковый шприц объемом 10 мл для стимуляции обоняния. Фильтровальную бумагу без запаха и без растворителя использовали в качестве контрольного стимула для тестирования механической стимуляции (воздух). Изоамилацетат и масляная кислота использовались в качестве условных раздражителей (CS + или CS-), в то время как транс-коричный альдегид служил в качестве контрольного запаха без каких-либо условных обстоятельств (награда или наказание).Запахи были выбраны на основе выбора поведения тараканов в предварительных испытаниях в Т-образном лабиринте, в которых не было обнаружено предпочтения между изоамилацетатом и масляной кислотой.
Изоамилацетат и масляная кислота использовались в качестве условных раздражителей (CS + или CS-), в то время как транс-коричный альдегид служил в качестве контрольного запаха без каких-либо условных обстоятельств (награда или наказание).Запахи были выбраны на основе выбора поведения тараканов в предварительных испытаниях в Т-образном лабиринте, в которых не было обнаружено предпочтения между изоамилацетатом и масляной кислотой.
Рисунок 1 . Экспериментальная дизайн. Эскиз (A), классической установки кондиционирования и (B) оперантной установки кондиционирования. (C) Представление одного тренировочного блока в классической парадигме кондиционирования. Каждый блок содержит воздушную затяжку (Воздух), вознагражденный запах CS + в сочетании с раствором сахара (R), наказанный запах CS- в сочетании с физиологическим раствором (P) и контрольный запах (Ctrl, коричный альдегид). (D) Временная последовательность условного стимула (CS, запах) и безусловного стимула (США, сахароза или физиологический раствор) в одном тренировочном испытании во время классического кондиционирования. (E) Протоколы оперантного кондиционирования (I) с пятью последовательными тренировочными попытками (C1 – C5) и тестом на удержание (RT) через 35 минут и (II) с тремя последовательными тренировочными попытками (C1 – C3) и тестом на удержание после 24 ч.
Для оперантного кондиционирования использовался гибкий лабиринт, изготовленный по индивидуальному заказу. Стены из поливинилхлорида позволяли легко очищать спиртом между разовыми испытаниями.Лабиринт был помещен на пластину земли, и различные части туннеля были объединены в Т-образный лабиринт (рис. 1B). Жалюзи позволяли закрывать стартовый и целевой боксы (20 см × 28 см × 4 см). Все эксперименты с Т-образным лабиринтом проводились при красном свете (рис. 1В).
Процедуры обучения и тестирования
Во-первых, мы установили новую классическую парадигму кондиционирования запряженного таракана, обучая животных проявлять определенные движения верхней челюсти и половых губ (ротового аппарата) в качестве условного ответного поведения. Мы назвали этот ответ реакцией верхней челюсти и половых губ (MLR). Прикасаясь к антеннам и ротовым частям раствором сахарозы, тараканы начинают быстро двигаться и расширять свои верхние челюсти и губы, самые центральные части ротового аппарата, чтобы потянуться к раствору и высосать его. Когда подается физиологический раствор, животные прикасаются к нему и пробуют его на вкус, не проглатывая, и демонстрируют явное поведение избегания (втягивание ротового аппарата). В каждом отдельном испытании появление или отсутствие MLR регистрировалось как двоичный ответ (0/1).Только если MLR наблюдался в течение первых 3 секунд появления запаха (до начала УЗИ, см. Рисунок 1C и описание ниже), это считалось условной реакцией. Эта новая парадигма классического кондиционирования тараканов аналогична парадигме реакции на удлинение хоботка, используемой в классическом кондиционировании медоносных пчел, впервые установленной Такеда (1961), а затем стандартизированной Биттерманом и др. (1983).
Мы назвали этот ответ реакцией верхней челюсти и половых губ (MLR). Прикасаясь к антеннам и ротовым частям раствором сахарозы, тараканы начинают быстро двигаться и расширять свои верхние челюсти и губы, самые центральные части ротового аппарата, чтобы потянуться к раствору и высосать его. Когда подается физиологический раствор, животные прикасаются к нему и пробуют его на вкус, не проглатывая, и демонстрируют явное поведение избегания (втягивание ротового аппарата). В каждом отдельном испытании появление или отсутствие MLR регистрировалось как двоичный ответ (0/1).Только если MLR наблюдался в течение первых 3 секунд появления запаха (до начала УЗИ, см. Рисунок 1C и описание ниже), это считалось условной реакцией. Эта новая парадигма классического кондиционирования тараканов аналогична парадигме реакции на удлинение хоботка, используемой в классическом кондиционировании медоносных пчел, впервые установленной Такеда (1961), а затем стандартизированной Биттерманом и др. (1983).
Для классического кондиционирования каждый блок тренировки состоял из (1) одной стимуляции простой фильтровальной бумагой без одоранта для проверки механической реакции на затяжку воздуха (Воздух), (2) одной парной презентации CS + (усиленный условный стимул) с 20% раствором сахарозы в качестве положительного подкрепления (безусловный стимул, УЗИ), (3) одна CS- презентация (наказываемый стимул) в сочетании с 20% физиологическим раствором в качестве отрицательного подкрепления, а также (4) одна стимуляция с контрольным запахом (коричный альдегид). , Ctrl), который не был связан с US (рис. 1C).В каждой презентации CS + или CS- соответствующий запах (CS) присутствовал в течение 15 с. Через три секунды после появления запаха УЗИ вводили путем прикосновения к верхнечелюстным пальцам сахарозы или физиологического раствора, и животным позволяли пить соответствующие растворы в течение 14 секунд (рис. 1D). В случае отрицательного подкрепления большинство животных не пили физиологический раствор добровольно, а были «вынуждены» пробовать соль во всех испытаниях, прикасаясь зубочисткой к ротовому аппарату. Мы провели все эксперименты в двух независимых группах, для которых идентичность запаха CS + и CS- была обратной.Для тестов на удерживание использовался тот же образец представления запаха, что и в испытаниях кондиционирования, за исключением того, что УЗИ не было представлено.
Мы провели все эксперименты в двух независимых группах, для которых идентичность запаха CS + и CS- была обратной.Для тестов на удерживание использовался тот же образец представления запаха, что и в испытаниях кондиционирования, за исключением того, что УЗИ не было представлено.
Было проведено три дифференциальных классических эксперимента по кондиционированию. В каждом блоке тренировочных испытаний два контрольных стимула (воздух, коричный альдегид) отделялись от двух CS с межстимульным интервалом (ISI) 45 с, тогда как ISI между CS + и CS- составляло 32 с. Первый эксперимент был разработан для исследования дифференциального обучения с фазой освоения, которая состояла из пяти блоков испытаний (каждый блок содержал одно представление воздуха, CS +, CS- и контрольную стимуляцию соответственно) с интервалом между блоками 10 минут.Через 10 мин был проведен тест на удерживание. Второй и третий эксперименты были разработаны для изучения сохранения памяти после дифференциального обучения в двух разных временных интервалах (1 час и 24 часа). Из-за продолжительности эксперимента для этих двух экспериментов использовались только три обучающих блока с интервалом между блоками 10 мин.
Для оперантного кондиционирования каждому таракану позволяли акклиматизироваться в стартовом боксе в течение 15 мин перед тренировкой. В начале тренировочного испытания заслонку открыли, и тараканам разрешили свободно ходить и входить в целевые ящики.При первом входе в один из ящиков-мишеней заслонка закрывалась и животное подвергалось 5-минутному световому воздействию (наказывающий раздражитель, УЗИ). Каждый раз, когда животное снова попадало в тот же ящик-мишень в последующем испытании, оно снова подвергалось легкому наказанию. Все животные, которые не начинали двигаться в течение первых 3 мин в двух последовательных попытках, исключались из эксперимента.
Были использованы две разные парадигмы оперантного кондиционирования. В первом эксперименте животные были обучены в пяти испытаниях с интервалом между испытаниями (ITI) 35 минут, а сохранение памяти проверялось через 35 минут. Во втором случае животные были обучены в трех испытаниях с ITI, равным 35 мин, и испытание на удержание было выполнено через 24 часа (рис. 1E).
Во втором случае животные были обучены в трех испытаниях с ITI, равным 35 мин, и испытание на удержание было выполнено через 24 часа (рис. 1E).
Статистика
Результаты были проанализированы с помощью Matlab R2019a (The MathWorks, Натик, Массачусетс, США) и IBM SPSS Statistics Version 25.0 (IBM Corp., Армонк, Нью-Йорк, США) и визуализированы с помощью Matlab R2019a и Photoshop (Adobe Inc., Сан-Хосе). , Калифорния, США).
Мы проанализировали спонтанные реакции на разные запахи у двух групп животных.Мы объединили поведенческую реакцию на представление запаха в первом тренировочном испытании и перед презентацией в УЗИ всех людей, которые лечились параллельно и в идентичных экспериментальных условиях. Для сравнения реакции на различные запахи использовались тесты хи-квадрат. Кроме того, мы рассчитали коэффициент Phi, чтобы проанализировать корреляцию между ответами на запах у разных людей.
Для статистического анализа классических экспериментов с кондиционированием мы применили три различных статистических теста.Во-первых, односторонний дисперсионный анализ был использован для проверки эволюции ответов на обучающих маршрутах. Во-вторых, для сравнения типа подкрепления (CS + и CS-) и типа подкрепления × пробное взаимодействие использовался двухфакторный дисперсионный анализ ANOVA с повторными измерениями. Хотя ANOVA обычно не допускается в случае дихотомических данных, таких как MLR, исследования Монте-Карло показали, что ANOVA можно использовать при определенных условиях (Lunney, 1970), которым все соответствуют эксперименты, описанные здесь. В-третьих, тесты χ 2 использовались для (1) сравнения ответов на CS + и CS- в данном испытании, (2) сравнения спонтанных ответов и тестов удержания, (3) сравнения последнего обучающего испытания и тесты удерживания и (4) сравнение различных тестов удерживания.
Для дальнейшего анализа классических экспериментов по кондиционированию мы объединили всех животных с одинаковым образцом кондиционирования независимо от запаха, который использовался как CS + или CS-. Чтобы проанализировать реакцию на CS +, мы исключили всех животных, показывающих спонтанные ответы на CS + в первом испытании. Для анализа ответов на CS- мы исключили всех животных, которые не ответили на него в первом испытании. Это обычная процедура для исключения спонтанно реагирующих животных и визуализации кривой обучения (Памир и др., 2011; Джурфа и Сандос, 2012).
Чтобы проанализировать реакцию на CS +, мы исключили всех животных, показывающих спонтанные ответы на CS + в первом испытании. Для анализа ответов на CS- мы исключили всех животных, которые не ответили на него в первом испытании. Это обычная процедура для исключения спонтанно реагирующих животных и визуализации кривой обучения (Памир и др., 2011; Джурфа и Сандос, 2012).
Для всех оперантных парадигм обусловливания решения в задачах с принудительным выбором двух вариантов были проанализированы с помощью биномиального теста, поскольку уровень вероятности случайного выбора одного из двух направлений составлял p = 0,5.
Анализ индивидуальности
Чтобы проанализировать индивидуальность обучающего поведения, мы следовали анализу Pamir et al. (2011, 2014). Для анализа в классической парадигме кондиционирования мы рассматривали только животных, которые не показали правильного ответа ни на CS +, ни на CS- в самом первом испытании и до того, как были представлены УЗИ.Две подгруппы были сформированы для тренировочных испытаний и тестовых испытаний в соответствии с определением, данным Pamir et al. (2011). Для любого данного испытания в первую подгруппу входили животные, которые проявляли правильное поведение в настоящем испытании и в предыдущем испытании (предыдущее правильное поведение, pC). Во вторую подгруппу вошли животные, показавшие правильное поведение в настоящем испытании, но не показавшие его в предыдущем испытании (предыдущее неправильное поведение, pI). Те же определения подгрупп были использованы для теста на удержание в отношении правильного или неправильного ответа во время заключительного обучающего испытания.Мы сравнили две подгруппы в каждом испытании и в тесте удержания с тестом × 2 с уровнем значимости α = 0,05. Две подгруппы представлены треугольниками, направленными вверх и вниз, соответственно. Закрашенные (открытые) символы указывают на то, что значимость не может быть установлена.
После анализа, проведенного Памиром и соавт. (2014), мы сформировали отдельные подгруппы животных, которые впервые показали правильное поведение во втором (или третьем) испытании, и отслеживали поведение подгрупп в последующих испытаниях кондиционирования и тесте на память.Это позволяет оценить устойчивость выражения правильного поведения в испытаниях и перенос поведенческого выражения во время обучения в тестовую ситуацию краткосрочной и долгосрочной памяти.
(2014), мы сформировали отдельные подгруппы животных, которые впервые показали правильное поведение во втором (или третьем) испытании, и отслеживали поведение подгрупп в последующих испытаниях кондиционирования и тесте на память.Это позволяет оценить устойчивость выражения правильного поведения в испытаниях и перенос поведенческого выражения во время обучения в тестовую ситуацию краткосрочной и долгосрочной памяти.
Наконец, чтобы проанализировать начало правильного поведения, мы вычислили для каждого испытания долю животных, которые впервые правильно ответили в этом испытании, а также долю животных, которые никогда не вели себя правильно (не учащиеся, Pamir et al. др., 2014).
Результаты
Спонтанная реакция на разные запахи
Сначала мы проанализировали спонтанные и наивные реакции на каждый запах (изоамилацетат, масляная кислота и коричный альдегид) во время самого первого эксперимента по кондиционированию до того, как был предъявлен усиливающий стимул (УЗ).На рисунке 2 показаны усредненные по группе реакции на все три запаха. Во всех экспериментах ок. 60% животных показали спонтанный MLR в присутствии изоамилацетата, который был значительно выше, чем для масляной кислоты и коричного альдегида для всех экспериментов ( × 2 : p <0,001). Количество спонтанных ответов на масляную кислоту и коричный альдегид значительно различается только в первом эксперименте ( × 2 : p = 0.01), у которых животные чаще спонтанно реагировали на масляную кислоту. Изоамилацетат является основным компонентом банановой смеси и, таким образом, тесно связан с пищей и привлекателен для тараканов (Lauprasert et al., 2006). Это, скорее всего, причина очень высокой скорости спонтанного ответа на изоамилацетат. Кроме того, мы обнаружили значительную положительную корреляцию между ответами на изоамилацетат и масляную кислоту в обоих экспериментах [Рисунок 2: (1) Φ = 0,0258, p = 0,016; (2) Φ = 0. 143, p = 0,017]. Однако не было значимой корреляции для других пар запахов, что могло быть связано с в целом низкой скоростью спонтанной реакции на коричный альдегид и масляную кислоту.
143, p = 0,017]. Однако не было значимой корреляции для других пар запахов, что могло быть связано с в целом низкой скоростью спонтанной реакции на коричный альдегид и масляную кислоту.
Рисунок 2 . Спонтанная реакция на изоамилацетат (ISO), масляную кислоту (BA) и коричный альдегид (CIN). Анализ MLR в первом испытании двух классических экспериментов по кондиционированию показывает, что ISO вызывает значительно больше спонтанных ответов, чем BA и CIN ( × 2 : p <0.001). Спонтанный ответ на БА только значительно выше по сравнению с КИН в первом эксперименте ( × 2 : p = 0,01).
Новая парадигма классического кондиционирования запахов
Мы установили новую парадигму классического кондиционирования запряженных тараканов (рис. 1А). Возникновение или отсутствие реакции верхней челюсти и половых губ (MLR, см. Раздел «Материалы и методы») регистрировали как поведение условной реакции (CR). В этом исследовании мы использовали разные протоколы для дифференциального обонятельного кондиционирования (рисунки 1C, D), чтобы исследовать экспрессию CR во время обучения и сохранения памяти в двух разных временных точках.
В первом протоколе мы проверили, способны ли тараканы ассоциировать запах с наградой или наказанием, в течение пяти последовательных тренировочных испытаний (интервал между испытаниями 10 минут) с последующим тестом на удержание (через 10 минут). Мы обучили две группы животных, для которых запахи изоамилацетата и масляной кислоты были представлены как CS + и CS- с обратными случайностями (рисунки 3A, B, соответственно). Два запаха не вызвали одинаковый уровень спонтанных реакций (см. Раздел «Спонтанная реакция на разные запахи»).Из-за высокой начальной спонтанной реакции на изоамилацетат средний уровень MLR был постоянно высоким и не увеличивался значительно в течение пяти испытаний, когда изоамилацетат использовался в качестве CS + (рис. 3A). Тем не менее, ответы на наказываемый запах (CS-, масляная кислота) значительно снизились (односторонний дисперсионный анализ: F 4, 260 = 4,23; p <0,002).
Рисунок 3 . Классическое обонятельное кондиционирование в пяти испытаниях с тестом на сохранение памяти через 10 мин.Закрашенные квадраты обозначают испытания, в которых была обнаружена значительная разница между CS + и CS- (тест × 2 ). Звездочки показывают, что ответы на условные стимулы (CS + и CS-) изменились в ходе испытаний (двухфакторный дисперсионный анализ, повторные измерения: лечение × пробное взаимодействие *: p <0,05; ***: p <0,001). (A) Когда изоамилацетат (ISO) был вознагражденным запахом (CS +, черный), средний ответ группы не увеличивался во время тренировки.Ответы на наказываемый запах BA (CS-, темно-серый) значительно снизились в течение испытаний (односторонний ANOVA: p = 0,002), и взаимодействие запах × испытание было значимым (ANOVA двухсторонних повторных измерений: p = 0,02) . (B) Когда BA был вознагражден (CS +, черный), ответы значительно увеличились, но не значительно (однофакторный дисперсионный анализ: p = 0,208). Ответы на ISO уменьшились, но опять же, незначительно (однофакторный дисперсионный анализ: p = 0,189). Взаимодействие запах × проба не было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p = 0.113). (C) При исключении всех животных, спонтанно реагирующих на запах, ответ на CS + значительно увеличился (односторонний ANOVA: p <0,001). Ответы на CS- уменьшились в течение испытаний, но незначительно (однофакторный дисперсионный анализ: p = 0,686). Взаимодействие лечение × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). (D) При исключении всех животных, не реагирующих спонтанно на наказываемый запах, количество животных, которые вели себя правильно в отношении CS- в ходе испытаний, значительно увеличилось (односторонний дисперсионный анализ: p <0. 001), тогда как ответ на CS + существенно не изменился в испытаниях (однофакторный дисперсионный анализ: p = 0,814). Взаимодействие лечение × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). Черные квадраты в (C, D) указывают на исключение животных, которые вели себя некорректно в первом испытании.
001), тогда как ответ на CS + существенно не изменился в испытаниях (однофакторный дисперсионный анализ: p = 0,814). Взаимодействие лечение × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). Черные квадраты в (C, D) указывают на исключение животных, которые вели себя некорректно в первом испытании.
Когда масляная кислота использовалась в качестве CS +, ответы показали тенденцию к увеличению в течение пяти тренировочных испытаний. В этом случае реакция на изоамилацетат как CS- немного снизилась; однако этот эффект не был значительным в течение пяти испытаний.Животные по-прежнему показали примерно 30% ответов на CS- в пятом испытании.
Ответы на контрольный запах коричного альдегида значительно снижались только тогда, когда изоамилацетат был CS + (односторонний дисперсионный анализ: F 4, 260 = 3,11; p = 0,016), но не изменился при использовании масляной кислоты в качестве CS +.
В целом, ответы на CS + и CS- значительно различались при вознаграждении изоамилацетата (двухфакторный дисперсионный анализ с повторными измерениями: F 4, 49 = 3.095; р = 0,02). Когда добавляли масляную кислоту, CS + и CS- существенно не различались в испытаниях (рис. 3B).
Для дальнейшего анализа мы объединили всех животных в соответствии с CS + и CS- и исключили тех, которые вели себя некорректно в первом испытании, соответственно. В обоих случаях правильное поведение значительно увеличивалось в ходе тренировочных испытаний [односторонний дисперсионный анализ: Рисунок 3C (CS +): F 4, 230 = 8,808; p <0,001; Рисунок 3D (CS-): F 4, 190 = 15.544; p <0,001]. Однако ни поведение CS- на рис. 3C, ни CS + на рис. 3D, ни поведение контрольного запаха существенно не изменились за время испытаний. Более того, взаимодействие между испытанием и лечением было значимым для CS + (ANOVA с двусторонними повторными измерениями: F 4, 43 = 12,156, p <0,001) и CS- (ANOVA с двусторонними повторными измерениями: F 4, 35 = 17,591, p <0,001), и в обоих случаях поведение в тестах на удерживание значительно отличалось друг от друга (рисунки 3C, D).
Кроме того, мы исключили животных, которые вели себя некорректно в отношении CS +, CS- или контроля. Соответственно, мы могли видеть, что эффект увеличения правильного поведения по сравнению с испытаниями был не только из-за исключения спонтанно реагирующих или не отвечающих животных (см. Дополнительный рисунок S1).
Выражение кратковременной и долговременной памяти после классической обработки
Чтобы проверить сохранение памяти после дифференциального классического кондиционирования в краткосрочной и долгосрочной перспективе, мы провели новый эксперимент.Группу тараканов дрессировали по-разному в течение трех последовательных испытаний. Затем группа была разделена пополам, и тесты на удержание были выполнены либо через 1 час после последнего тренировочного испытания, либо через 24 часа после него. На рисунке 4 мы показываем тренировочные испытания как неразделенные группы, но статистический анализ, включающий тренировочные испытания, проводился с разделенными группами, которые показаны на дополнительном рисунке S2. Эксперимент был повторен с обратной случайностью запахов. Реакция на механическое воздействие воздуха (только фильтровальная бумага) не менялась и всегда оставалась ниже 1.5%.
Рисунок 4 . Классическое обонятельное кондиционирование в трех испытаниях с тестами на сохранение памяти через 1 и 24 часа. Закрашенные квадраты обозначают испытания, в которых была обнаружена значительная разница между CS + и CS- (тест × 2 ). Звездочки указывают на значимое взаимодействие лечение × испытание (***: p <0,001). Буквы над столбиками указывают на существенные различия между тестами на удерживание ( × 2 ). (A) Когда изоамилацетат (ISO) был вознагражден (CS +, черный) и масляная кислота (BA) наказана (CS-, темно-серый), ответы значительно снизились для CS- (односторонний ANOVA: p < 0.001), но не изменилось для CS + (односторонний дисперсионный анализ: p = 0,155). В целом, взаимодействие запах × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). (B) Когда масляная кислота была вознаграждена (CS +, черный), а изоамилацетат был наказан (CS-, темно-серый), ответы на CS + увеличились, однако, незначительно (односторонний ANOVA: p = 0,06) , но значительно снизился для CS- (односторонний дисперсионный анализ: p = 0,002). Взаимодействие запах × испытание снова было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0.001). (C) При исключении всех животных, спонтанно реагирующих на запах, ответ на CS + значительно увеличился (односторонний ANOVA: p <0,001). Ответ на CS- значительно снизился в течение испытаний (однофакторный дисперсионный анализ: p = 0,029). Взаимодействие лечение × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). (D) При исключении всех животных, не реагирующих спонтанно на наказываемый запах, количество животных, которые вели себя правильно в отношении CS- в ходе испытаний, значительно увеличилось (односторонний дисперсионный анализ: p <0.001). Ответ на CS + существенно не изменился в испытаниях (однофакторный дисперсионный анализ: p = 0,38). Взаимодействие лечение × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). Черный квадрат в (C, D) указывает на исключение животных, которые вели себя некорректно в первом испытании.
В целом, взаимодействие запах × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). (B) Когда масляная кислота была вознаграждена (CS +, черный), а изоамилацетат был наказан (CS-, темно-серый), ответы на CS + увеличились, однако, незначительно (односторонний ANOVA: p = 0,06) , но значительно снизился для CS- (односторонний дисперсионный анализ: p = 0,002). Взаимодействие запах × испытание снова было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0.001). (C) При исключении всех животных, спонтанно реагирующих на запах, ответ на CS + значительно увеличился (односторонний ANOVA: p <0,001). Ответ на CS- значительно снизился в течение испытаний (однофакторный дисперсионный анализ: p = 0,029). Взаимодействие лечение × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). (D) При исключении всех животных, не реагирующих спонтанно на наказываемый запах, количество животных, которые вели себя правильно в отношении CS- в ходе испытаний, значительно увеличилось (односторонний дисперсионный анализ: p <0.001). Ответ на CS + существенно не изменился в испытаниях (однофакторный дисперсионный анализ: p = 0,38). Взаимодействие лечение × испытание было значимым (двухфакторный дисперсионный анализ с повторными измерениями: p <0,001). Черный квадрат в (C, D) указывает на исключение животных, которые вели себя некорректно в первом испытании.
В целом, ответы на CS + значительно отличались от CS- в трех тренировочных испытаниях в обеих группах (двухфакторный дисперсионный анализ с повторными измерениями: Рисунок 4A: F 2, 183 = 9.266, p <0,001; Фигура 4В: F 2, 91 = 13,016; р <0,001). Ответы на CS- значительно снизились в обоих экспериментах (односторонний дисперсионный анализ: Рисунок 4A: F 2, 552 = 7,181; p <0,001; Рисунок 4B: F 2, 276 = 3,291; p = 0,002), в то время как ответы на CS + существенно не увеличились. Ответы на контрольный запах коричного альдегида значительно снижались только при использовании масляной кислоты в качестве CS + (односторонний дисперсионный анализ: F 2, 276 = 3.291; р = 0,039).
Ответы на контрольный запах коричного альдегида значительно снижались только при использовании масляной кислоты в качестве CS + (односторонний дисперсионный анализ: F 2, 276 = 3.291; р = 0,039).
В течение 1 часа испытуемые животные, которые получали изоамилацетат в виде CS +, сохраняли повышенный уровень ответа, так как результативность усредненной по группе, таким образом, тест удержания через 1 час существенно не отличался от уровня ответа в последнем тренировочном испытании (рис. 2 : p = 0,143). Интересно, что уровень ответа на изоамилацетат был значительно выше в тесте на 24-часовое удерживание по сравнению с 1-часовым тестом на удержание ( × 2 : p <0.001), а также по сравнению с уровнем ответа в конце обучения ( × 2 : p <0,001) (дополнительная таблица S1). Когда масляная кислота была вознагражденным запахом (рис. 6В), уровни реакции на CS + в обоих тестах памяти (через 1 и 24 часа) не отличались друг от друга ( × 2 : p = 0,475), ни от уровня ответа в конце обучения ( χ 2 : 1 ч: р = 0,826; 24 ч: р = 0.149). Уровень ответа на CS- возобновил первоначальные высокие уровни спонтанного ответа на изоамилацетат в течение 1 и 24 часов удерживания. Все результаты приведены в дополнительной таблице S2.
Для следующего этапа анализа были исключены животные, которые некорректно вели себя в первом испытании, и как для CS +, так и для CS- процент правильно ведущих себя животных увеличился (Рисунки 4C, D, однофакторный дисперсионный анализ ANOVA: CS +: F 2, 537 = 33,537; p <0,001; CS-: F 2, 306 = 43.027; р <0,001). Единственным другим значимым эффектом было уменьшение правильного поведения по отношению к CS-, когда все правильно ответившие животные были исключены (односторонний ANOVA: F 2, 537 = 3,569; p = 0,029). Более того, взаимодействие между испытанием и лечением было значительным в обоих случаях (двухфакторный дисперсионный анализ с повторными измерениями: Рисунок 4C: F 2, 178 = 46,719; p <0,001; Рисунок 4D: F 2, 101 = 39.158; р <0,001).
Более того, взаимодействие между испытанием и лечением было значительным в обоих случаях (двухфакторный дисперсионный анализ с повторными измерениями: Рисунок 4C: F 2, 178 = 46,719; p <0,001; Рисунок 4D: F 2, 101 = 39.158; р <0,001).
При исключении всех спонтанно реагирующих животных результативность теста удержания оставалась на том же уровне, что и в конце обучения. Однако увеличение показателя удерживания CS + через 24 часа было значительным ( × 2 : p <0,001). После исключения неспонтанных респондентов результаты всех тестов удержания оставались аналогичными по сравнению с третьим испытанием обучения. Однако процент правильного поведения животных по отношению к CS- снизился через 1 час ( × 2 : p <0.001) и 24 часа ( χ 2 : p <0,001).
Эффективность индивидуального обучения во время классической подготовки
Чтобы проверить, существуют ли различия в успеваемости у отдельных животных, мы следовали анализу, предложенному в Pamir et al. (2011, 2014). Каждая из двух групп, обученных в пяти пробных классических экспериментах по кондиционированию (рисунки 3C, D), была разделена на две подгруппы (см. Раздел «Материалы и методы»): (1) люди, которые вели себя правильно в двух последовательных испытаниях (предыдущее правильное поведение , pC) и (2) люди, которые не вели себя правильно в предыдущем испытании, но начали вести себя правильно в настоящем испытании (предыдущее неправильное поведение, pI).Предыдущие животные с правильным поведением всегда демонстрировали более высокий уровень правильного поведения, чем среднее правильное поведение среди всех животных (рис. 5, красные треугольники, направленные вверх), в то время как предыдущие животные с неправильным поведением всегда демонстрировали более низкое правильное поведение (рис. 5, красные треугольники вниз).
Рисунок 5 . Индивидуальные способности к обучению классическому обонятельному кондиционированию. Данные для изоамилацетата в виде CS + и масляной кислоты в качестве CS + были объединены, аналогично данные для изоамилацетата в качестве CS- и масляной кислоты в качестве CS- были объединены.Правильное обучение представлено (A), , увеличивающими ответами на вознагражденный запах в случае CS + и (C), , увеличивающими подавление ответа на наказываемый запах в случае CS-. (A, C) Вверху указывающими треугольниками изображены животные, которые вели себя правильно в соответствующем испытании и уже показали правильное поведение в предыдущем испытании (ПК). Треугольники, указывающие вниз, изображают животных, которые вели себя правильно в соответствующем испытании, но не вели себя в предыдущем испытании (pI).Заштрихованные треугольники указывают на достоверное различие подгрупп в соответствующих испытаниях, пустые треугольники представляют подгруппы, которые существенно не различались ( × 2 ; α = 0,05). Животные, правильно ведущие себя во втором испытании, т. Е. После единственного тренировочного испытания (темно-серый; CS +: N = 15; CS-: N = 16), показали стабильно высокие вероятности правильных ответов в последующих испытаниях (см. Pamir et al. др., 2014). Аналогичным образом, высокие показатели ответа наблюдались у животных, впервые правильно ответивших в третьем испытании (светло-серый; CS +: N = 11; CS-: N = 11).Черные квадраты указывают на исключение животных, которые в первом испытании вели себя некорректно. (B, D) Доля животных, впервые ведущих себя правильно в указанном испытании. Большой процент животных проявил правильное поведение уже после однократной тренировки. Значительная часть животных никогда не вела себя правильно (черные полосы).
Затем мы проанализировали поведение тех животных, которые показали свое первое правильное поведение уже во втором испытании (т.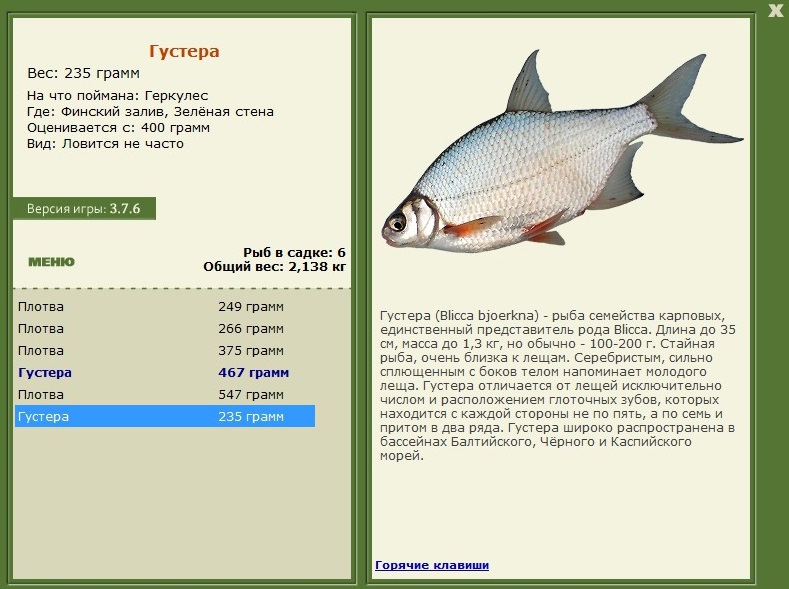 е., после всего лишь одной пары CS и US) и обнаруживают, что эта подгруппа показывала стабильно высокие показатели правильных ответов во всех испытаниях и во время удержания с оценками удержания выше 90%, как для CS +, так и для CS- (темно-серые кривые на рисунках). 5А, В). Лица, которые начали правильно реагировать в третьем испытании (после двух пар CS и US, светло-серые кривые на рисунках 5A, C) показали более низкие уровни правильного ответа, чем те животные, которые начали в первом испытании, но, тем не менее, они были сравнительно сопоставимыми. высокий, учитывая тот факт, что средние уровни отклика (синяя и зеленая кривая на рисунках 5A, C, соответственно) включали также группу с высокими характеристиками (темно-серые кривые).Это указывает на то, что быстро обучающиеся также хорошо обучаются, и соответствует предыдущим результатам, полученным на пчелах (Памир и др., 2014).
е., после всего лишь одной пары CS и US) и обнаруживают, что эта подгруппа показывала стабильно высокие показатели правильных ответов во всех испытаниях и во время удержания с оценками удержания выше 90%, как для CS +, так и для CS- (темно-серые кривые на рисунках). 5А, В). Лица, которые начали правильно реагировать в третьем испытании (после двух пар CS и US, светло-серые кривые на рисунках 5A, C) показали более низкие уровни правильного ответа, чем те животные, которые начали в первом испытании, но, тем не менее, они были сравнительно сопоставимыми. высокий, учитывая тот факт, что средние уровни отклика (синяя и зеленая кривая на рисунках 5A, C, соответственно) включали также группу с высокими характеристиками (темно-серые кривые).Это указывает на то, что быстро обучающиеся также хорошо обучаются, и соответствует предыдущим результатам, полученным на пчелах (Памир и др., 2014).
На гистограммах на рисунках 5B, D мы подсчитали для каждого испытания отдельно, сколько животных ответили правильно впервые в этом испытании. Из всех животных, которые продемонстрировали способность к обучению, большинство из них показали правильную реакцию после однократного испытания. Вторая по величине группа впервые вела себя правильно после двух проб по кондиционированию. Однако значительная часть животных никогда не вела себя правильно (черные полосы на рисунках 5B, D), и эта группа больше для правильного поведения по отношению к CS +.
Индивидуальность в оперантном обучении
Затем мы проверили обучение, сохранение памяти и индивидуальные различия в задаче оперантного кондиционирования. Тараканов дрессировали избегать наказания и проверяли на память в течение 24 часов. Для этого мы разработали принудительную парадигму с двумя вариантами выбора, в которой отдельный таракан помещается в Т-образный лабиринт во время повторных тренировочных испытаний (рис. 1В, см. Материалы и методы). В каждом испытании таракану разрешалось выбирать одну из рук и входить в поле-мишень. В первом испытании, независимо от выбора животного, оно испытывало неприятный раздражитель ярким светом. Каждый раз, когда животное выбирало одну и ту же сторону в последующих испытаниях, вызывался один и тот же отвращающий стимул. Таким образом, обучение выражалось в избегании той стороны (левой или правой), которая приводила к наказанию ярким световым стимулом.
В первом испытании, независимо от выбора животного, оно испытывало неприятный раздражитель ярким светом. Каждый раз, когда животное выбирало одну и ту же сторону в последующих испытаниях, вызывался один и тот же отвращающий стимул. Таким образом, обучение выражалось в избегании той стороны (левой или правой), которая приводила к наказанию ярким световым стимулом.
В первом эксперименте животных обучали для пяти последовательных испытаний, а сохранение кратковременной памяти проверяли через 35 минут после последнего испытания. Во втором эксперименте животные были обучены для трех испытаний, и через 24 часа был проведен тест на долгосрочное удержание (рис. 1E).Животные в первой группе значительно научились избегать наказываемой стороны начиная с третьего испытания (биномиальные тесты: p <0,01). Животные показали правильную память для наказанной стороны в тесте на удержание через 35 минут (фигура 6А). Тараканы из второй группы не показали значимой обучаемости после двух тренировочных испытаний. Однако память о правильной стороне была выражена в 24-часовом тесте на сохранение (биномиальный тест: p = 0,014, рис. 6В).
Рисунок 6 .Общая групповая производительность и индивидуальный выбор во время оперантной задачи кондиционирования в Т-образном лабиринте (A) в течение пяти последовательных испытаний и 35-минутного теста удержания (RT). Из третьего испытания на тараканах значительно чаще выбирали правильное направление (**: p <0,01; ***: p <0,001). (B) Оперантное кондиционирование в трех испытаниях и тест на удерживание через 24 часа. Однако на этот раз тараканы не обучались во время обучения, а выбрали значительно более правильное направление в тесте на 24-часовое удержание (*: p <0.05). Треугольники, указывающие вверх, изображают животных, которые выбрали правильное направление в двух последовательных испытаниях (ПК). Треугольники, направленные вниз, изображают животных, которые выбрали правильное направление в настоящем испытании, но не выбрали его в предыдущем испытании (pI). Заштрихованные треугольники достоверно отличаются друг от друга в рамках эксперимента, а пустые – нет ( χ 2 ; α = 0,05).
Заштрихованные треугольники достоверно отличаются друг от друга в рамках эксперимента, а пустые – нет ( χ 2 ; α = 0,05).
Для изучения индивидуальных различий в этих оперантных задачах обучения и запоминания животных снова разделили на две подгруппы.В эксперименте с кратковременной памятью животные в подгруппе, показавшие правильное поведение в двух последовательных испытаниях (pC), всегда показывали лучшие результаты, чем в среднем по группе, в то время как животные в подгруппе pI постоянно показывали меньше правильных выборов в настоящем испытании. Поведенческий выбор предыдущих правильно решающих животных (pC) и предыдущих неправильно решающих животных (pI) значительно различались в третьем и четвертом испытаниях во время обучения ( × 2 : p <0,05).Эта разница не была значимой ни в пятом обучающем испытании, ни в тесте на удержание через 35 минут ( × 2 : испытание 3: p = 0,161; испытание 4: p = 0,186, рис. 6А).
В эксперименте с долговременной памятью две подгруппы (pC и pI) снова значительно различались после двух тренировочных испытаний. Во время 24-часового теста на удержание подгруппа животных, показавших правильное решение в последнем тренировочном испытании, значительно превосходила тех животных, которые приняли неправильное решение в последнем тренировочном испытании (Рисунок 6B: × 2 : p <0.05).
Обсуждение
В настоящей работе мы показываем, что взрослый американский таракан, Periplaneta americana , может решать классические обонятельные и оперантные задачи пространственного кондиционирования. В обоих случаях животные могли научиться устанавливать условную реакцию на поощренный стимул (CS +) и уменьшать свою реакцию на наказываемый запах (CS-), несмотря на то, что в классической задаче кондиционирования эти два запаха не были одинаковыми. важно для животных (высокая спонтанная реакция на изоамилацетат). В целом, тренировка привела к успешному проявлению кратковременной и долговременной памяти (через 24 часа) в обоих упражнениях на кондиционирование. Мы также показываем, что тараканы выражают индивидуальность в их обучении и памяти при классическом и оперантном обусловливании.
В целом, тренировка привела к успешному проявлению кратковременной и долговременной памяти (через 24 часа) в обоих упражнениях на кондиционирование. Мы также показываем, что тараканы выражают индивидуальность в их обучении и памяти при классическом и оперантном обусловливании.
Классическая обонятельная обработка тараканов
Для настоящего исследования мы установили новую классическую парадигму обусловливания у запряженных тараканов, которая позволяет наблюдать проявление (или невыражение) дискретного условного ответного поведения, ответа m подмышечной впадины l abia r MLR) во время обучения и сохранения памяти.Развитие этой парадигмы было вдохновлено очень успешной парадигмой рефлекса удлинения хоботка (PER) у медоносной пчелы (например, Kuwabara, 1957; Takeda, 1961; Bitterman et al., 1983; Giurfa and Sandoz, 2012; Menzel, 2012). при котором расширение (или нерасширение) хоботка наблюдается как дискретная условная реакция.
В ряде предыдущих исследований изучалась классическая обусловленность таракана с использованием условий обучения и тестирования, которые в корне отличаются от нашей парадигмы MLR.В исследованиях Watanabe et al. (2003), Sato et al. (2006) и Liu and Sakuma (2013), у немецких тараканов необузданных тараканов помещали в цилиндрическую камеру во время повторных испытаний кондиционирования с одним запахом в сочетании с сахарозной наградой (CS +) и вторым запахом в сочетании с солевым наказанием (CS- ). Sato et al. (2006) смогли доказать, что помимо простого обучения обонятельной дискриминации, тараканы демонстрируют отличные способности к обучению в парадигме создания событий, в которой визуальный контекст определяет случайность между обонятельными CS (обусловливающими стимулами) и вкусовыми US (безусловными стимулами).Watanabe et al. (2003) распространили свой классический протокол кондиционирования на необузданных тараканах на запряженных тараканов, которые впоследствии были протестированы в условиях свободного движения на тестовой арене, где они могли выбирать между двумя ранее кондиционированными запахами. Эта парадигма, однако, не установила четкой условной реакции, наблюдаемой во время обучения, и, таким образом, выражение поведения условной реакции доступно только во время сохранения памяти и в условиях, отличных от обучения.Классическое кондиционирование приводит к усилению реакции нейронов слюны на запах, связанный с вознаграждением сахарозы у таракана (Watanabe and Mizunami, 2006). После дифференцированного кондиционирования, когда один запах сочетается с сахарозой, а другой запах – без вознаграждения, связанный с сахарозой запах вызывал увеличение уровня слюноотделения, но только представленный запах этого не делал, что доказывает классическое кондиционирование слюноотделения у тараканов (Watanabe and Mizunami, 2007). ). Классическая обусловленность слюноотделения была впервые показана столетие назад Павловым в его знаменитых экспериментах на собаках (Павлов, 1927).В дальнейшем сдерживаемые тараканы использовались для изучения пространственного (например, Kwon et al., 2004) или зрительно-обонятельного ассоциативного обучения и памяти (например, Lent and Kwon, 2004; Pintér et al., 2005; Lent et al., 2007) количественная оценка реакции проекции антенн (APR) животных, привязанных в середине арены (Pomaville and Lent, 2018). APR основан на наблюдении, что двигательные действия антенн могут быть вызваны различными способами, включая обонятельные, тактильные и зрительные стимулы (например,, Menzel et al., 1994; Erber et al., 1997). Кондиционирование APR состоит в количественной оценке направленных движений антенн в направлении награжденного визуального стимула, и было вдохновлено оперантным условием пчел вытягивать свои антенны к цели, чтобы получить вознаграждение (например, Menzel et al., 1994; Kisch and Эрбер, 1999). Преимущество обучения иммобилизованных насекомых обеспечивает мощную технику для изучения нейронной основы (например, с использованием нейрофизиологических и фармакологических методов) обучения и памяти в более простой нервной системе по сравнению с позвоночными.
Эта парадигма, однако, не установила четкой условной реакции, наблюдаемой во время обучения, и, таким образом, выражение поведения условной реакции доступно только во время сохранения памяти и в условиях, отличных от обучения.Классическое кондиционирование приводит к усилению реакции нейронов слюны на запах, связанный с вознаграждением сахарозы у таракана (Watanabe and Mizunami, 2006). После дифференцированного кондиционирования, когда один запах сочетается с сахарозой, а другой запах – без вознаграждения, связанный с сахарозой запах вызывал увеличение уровня слюноотделения, но только представленный запах этого не делал, что доказывает классическое кондиционирование слюноотделения у тараканов (Watanabe and Mizunami, 2007). ). Классическая обусловленность слюноотделения была впервые показана столетие назад Павловым в его знаменитых экспериментах на собаках (Павлов, 1927).В дальнейшем сдерживаемые тараканы использовались для изучения пространственного (например, Kwon et al., 2004) или зрительно-обонятельного ассоциативного обучения и памяти (например, Lent and Kwon, 2004; Pintér et al., 2005; Lent et al., 2007) количественная оценка реакции проекции антенн (APR) животных, привязанных в середине арены (Pomaville and Lent, 2018). APR основан на наблюдении, что двигательные действия антенн могут быть вызваны различными способами, включая обонятельные, тактильные и зрительные стимулы (например,, Menzel et al., 1994; Erber et al., 1997). Кондиционирование APR состоит в количественной оценке направленных движений антенн в направлении награжденного визуального стимула, и было вдохновлено оперантным условием пчел вытягивать свои антенны к цели, чтобы получить вознаграждение (например, Menzel et al., 1994; Kisch and Эрбер, 1999). Преимущество обучения иммобилизованных насекомых обеспечивает мощную технику для изучения нейронной основы (например, с использованием нейрофизиологических и фармакологических методов) обучения и памяти в более простой нервной системе по сравнению с позвоночными.
Первоначальное ответное поведение во время классического кондиционирования
Стимулы, используемые для изучения обонятельного обучения и памяти у насекомых, в основном используют запахи, которые имеют отношение к естественному контексту, такие как коммуникационные сигналы (например, феромоны) или запахи, связанные с едой. Изоамилацетат представляет собой наиболее заметное соединение банановой смеси и воспринимается как запах банана (Schubert et al., 2014). Этот запах явно связан с пищей и поэтому очень привлекателен для тараканов (Lauprasert et al., 2006). Это, вероятно, объясняет, почему в наших экспериментах по кондиционированию обоняния мы наблюдали высокий уровень первоначальной реакции на изоамилацетат в первом испытании (рис. 2). Следовательно, было трудно наблюдать обучение (то есть повышение уровней условной реакции), когда этот запах сочетался с вознаграждением сахарозой (CS +), поскольку уровни ответа были постоянно высокими (> ~ 60%) с первого испытания и на протяжении всего обучения (рис. 3А). Однако в тесте на 24-часовое удержание MLR для изоамилацетата был значительно увеличен по сравнению с ответом в последнем тренировочном испытании (рис. 4A), что указывает на установление долговременной памяти.Когда изоамилацетат сочетался с солевым наказанием (CS-), животные научились подавлять свои реакции во время обучения (рис. 4B). Первоначальная реакция на масляную кислоту была значительно ниже (~ 30%) в начале тренировки во всех случаях (рис. 2). Когда он был связан с сахаром, ответы увеличивались, но никогда не превышали 50% даже после пяти тренировочных проб. Спонтанные реакции на масляную кислоту полностью устранялись во время тренировки и сохранения памяти в сочетании с наказанием (рис. 3B). В итоге два запаха, использованные в нашем исследовании, не были одинаково привлекательными для животных.
Оперантное пространственное кондиционирование у тараканов
Часто используемая установка для оперантного кондиционирования представляет собой Y- или Т-образный лабиринт, который широко используется для изучения оперантного обучения и принятия решений у грызунов.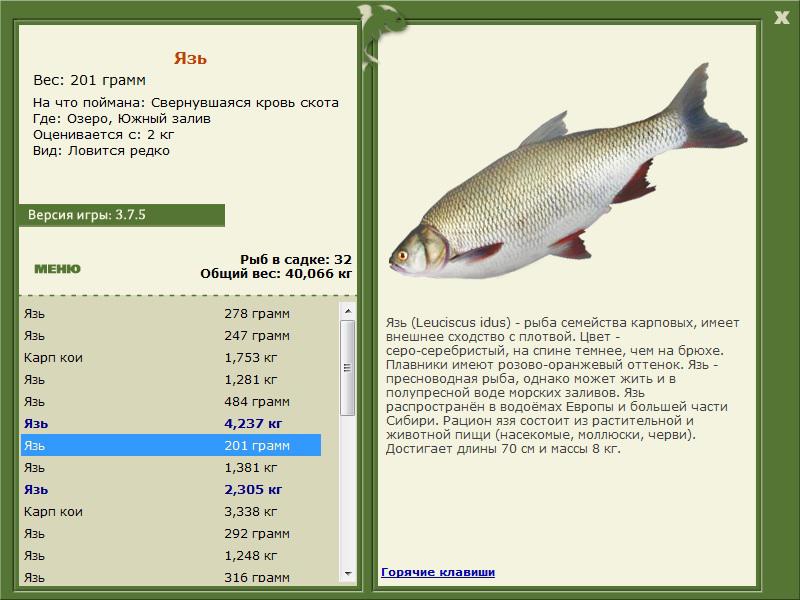 Эксперименты с Т- или Y-лабиринтом (двойной выбор) на беспозвоночных широко использовались для изучения визуального или обонятельного абсолютного и дифференциального обучения у свободно летающих пчел (например, для обзора: Srinivasan et al., 1998; Giurfa et al., 1999 , 2001; Avarguès-Weber et al., 2011; Nouvian, Galizia, 2019), у муравьев (e.г., Dupuy et al., 2006; Camlitepe, Aksoy, 2010) и ос (Hoedjes et al., 2012). У тараканов оперантное обучение неоднократно изучалось на открытых аренах (например, Balderrama, 1980; Sakura and Mizunami, 2001; Sakura et al., 2002). Первая работа по оперантному кондиционированию тараканов была проведена Балдеррамой (Balderrama, 1980), который обучал свободно движущихся тараканов индивидуально в простой тренировочной камере ассоциировать два искусственных запаха с растворами сахарозы и солевого раствора, соответственно, и тестировал дискриминирующую способность к обучению путем измерения запаха. предпочтение до и после тренировки.Спонтанное предпочтение одного из запахов перед тренировкой можно было изменить уже за одно испытание, и удержание длилось до 7 дней. На сегодняшний день существует только два исследования, в которых тараканов бросали вызов тараканам в задачах Т-образного лабиринта: первое проверяло влияние феромонов фекалий на направленную ориентацию (Bell et al., 1973), а второе исследовало влияние лекарств, ингибирующих синтез белка, на обучение. и удержание дрессировкой животных, чтобы избежать удара с одной из сторон (Barraco et al., 1981).Наша причина для реализации парадигмы оперантного обучения в Т-лабиринте заключалась в том, чтобы установить принудительный двоичный выбор, который может быть проанализирован во время сбора данных и сохранения памяти в рамках определенного дизайна исследования. Электрический шок, использованный в качестве наказывающего стимула в предыдущем исследовании Barraco et al. (1981) кажется довольно неестественным отвращающим стимулом, который вряд ли может появиться в природе. Мы решили использовать яркий свет в качестве отрицательного подкрепления, поскольку тараканы, естественно, избегают яркого света и ищут убежища в темноте (Turner, 1912).
Эксперименты с Т- или Y-лабиринтом (двойной выбор) на беспозвоночных широко использовались для изучения визуального или обонятельного абсолютного и дифференциального обучения у свободно летающих пчел (например, для обзора: Srinivasan et al., 1998; Giurfa et al., 1999 , 2001; Avarguès-Weber et al., 2011; Nouvian, Galizia, 2019), у муравьев (e.г., Dupuy et al., 2006; Camlitepe, Aksoy, 2010) и ос (Hoedjes et al., 2012). У тараканов оперантное обучение неоднократно изучалось на открытых аренах (например, Balderrama, 1980; Sakura and Mizunami, 2001; Sakura et al., 2002). Первая работа по оперантному кондиционированию тараканов была проведена Балдеррамой (Balderrama, 1980), который обучал свободно движущихся тараканов индивидуально в простой тренировочной камере ассоциировать два искусственных запаха с растворами сахарозы и солевого раствора, соответственно, и тестировал дискриминирующую способность к обучению путем измерения запаха. предпочтение до и после тренировки.Спонтанное предпочтение одного из запахов перед тренировкой можно было изменить уже за одно испытание, и удержание длилось до 7 дней. На сегодняшний день существует только два исследования, в которых тараканов бросали вызов тараканам в задачах Т-образного лабиринта: первое проверяло влияние феромонов фекалий на направленную ориентацию (Bell et al., 1973), а второе исследовало влияние лекарств, ингибирующих синтез белка, на обучение. и удержание дрессировкой животных, чтобы избежать удара с одной из сторон (Barraco et al., 1981).Наша причина для реализации парадигмы оперантного обучения в Т-лабиринте заключалась в том, чтобы установить принудительный двоичный выбор, который может быть проанализирован во время сбора данных и сохранения памяти в рамках определенного дизайна исследования. Электрический шок, использованный в качестве наказывающего стимула в предыдущем исследовании Barraco et al. (1981) кажется довольно неестественным отвращающим стимулом, который вряд ли может появиться в природе. Мы решили использовать яркий свет в качестве отрицательного подкрепления, поскольку тараканы, естественно, избегают яркого света и ищут убежища в темноте (Turner, 1912).
Тараканы стали избегать стороны, которую наказали после нескольких испытаний. Однако результаты обучения в двух экспериментах были разными. Предыдущие исследования пришли к выводу, что тараканы проявляют непредсказуемое поисковое поведение (Balderrama, 1980). Точно так же мы могли наблюдать разные черты поведения, которые частично могут лежать в основе вариативности поведения выбора. Например, некоторые тараканы демонстрируют активное исследовательское поведение, возможно, в поисках выхода из лабиринта, и они, похоже, не особо заботятся о подкрепляющем стимуле, в то время как другие оставались почти неподвижными на протяжении всего испытания и мало двигались.Карательный эффект света ограничен, потому что он не причиняет вреда животному. Таким образом, они могут привыкнуть к отталкивающему световому раздражителю. Эксперименты с Т-образным лабиринтом, проведенные Barraco et al. (1981) использование электрического шока в качестве отрицательного подкрепления привело к удивительно высоким показателям правильности выбора. Однако сильный свет кажется отталкивающим для большинства тараканов, поскольку они обычно пытаются спрятаться в темном месте, когда, например, в комнате включен свет. В будущих экспериментах мы хотим исследовать, может ли парадигма оперантного аппетита привести к более высокому уровню правильного выбора у тараканов.
Индивидуальное поведенческое выражение обучения и памяти
Наш подход к изучению индивидуальности в успеваемости во время классического кондиционирования был вдохновлен двумя предыдущими исследованиями Pamir et al. (2011, 2014), которые исследовали большое количество наборов данных по классическому условию аппетита у медоносных пчел. В этих исследованиях авторы смогли извлечь из огромного количества данных, что медоносные пчелы проявляют индивидуальное обучающее поведение и что группу животных можно разделить как минимум на две подгруппы: учащихся и не учащихся.Оба исследования, проведенные Памиром и его коллегами, изучали поведенческое обучение и выражение памяти только в отношении CS +. Мы расширили их анализ, включая поведенческое обучение и выражение памяти, в сторону CS- (рис. 5C, D).
Мы расширили их анализ, включая поведенческое обучение и выражение памяти, в сторону CS- (рис. 5C, D).
После исключения особей, которые не ответили правильно в первом испытании, как в случае медоносной пчелы (Памир и др., 2014), большая часть животных (> 35%) никогда не показывала правильного поведения в отношении запаха CS + ни в одном из обучающие испытания или тест на удержание (Рисунки 5B, D).Этих животных можно считать не обучающимися. Принимая во внимание только тех животных, которые хотя бы один раз проявили правильное условное поведение во время тренировки, мы обнаруживаем, что эти животные впервые проявили такое поведение после в среднем 1,7 испытаний кондиционирования по отношению к CS + и после в среднем 1,8 испытаний кондиционирования по отношению к CS. -. Действительно, самая большая фракция (50%) отвечающих животных впервые показала правильное условное ответное поведение уже после однократного испытания кондиционирования (однократное обучение) как в отношении CS +, так и CS-.Фактически, 86,6% учащихся показали первое правильное поведение по отношению к CS + или CS- уже после первого или второго испытания на кондиционирование, что указывает на быстрое обучение после одного или двух испытаний. Эти числа близко совпадают с данными, полученными для пчел, где обычно ~ 50% особей в группе медоносных пчел показали условный ответ после одного тренировочного испытания (Pamir et al., 2014). Более того, правильное выражение усвоенного поведения у быстро обучающихся является удивительно стабильным, что можно увидеть, проследив за CR-поведением в подгруппе тараканов, которое показало правильное поведение после однократного испытания (темно-серая кривая на рисунках 5A, C). ).Если посмотреть на сохранение кратковременной памяти у этих животных (рисунки 5A, C), 93,3 и 93,8% показали правильное поведение во время теста на CS + и CS-, соответственно. И наоборот, из фракции животных, которые показали правильное поведение при сохранении кратковременной памяти для CS + и CS-, 95 и 82,8%, соответственно, были быстро обучающимися индивидуумами, выражавшими правильное поведение после одного или двух тренировочных проб. Аналогичным образом Pamir et al. (2014) сообщили, что пчелы, которые ответили раньше, показали более высокую долговременную память, чем те, кто ответил позже.Взятые вместе, наши результаты показывают, что (1) отдельные тараканы способны эффективно учиться только в течение одного или двух испытаний кондиционирования, и (2) быстро обучающиеся также являются хорошими учениками, которые устойчиво выражают правильное поведение на протяжении всего сеанса обучения и достигают очень высокого удержания. оценки.
Аналогичным образом Pamir et al. (2014) сообщили, что пчелы, которые ответили раньше, показали более высокую долговременную память, чем те, кто ответил позже.Взятые вместе, наши результаты показывают, что (1) отдельные тараканы способны эффективно учиться только в течение одного или двух испытаний кондиционирования, и (2) быстро обучающиеся также являются хорошими учениками, которые устойчиво выражают правильное поведение на протяжении всего сеанса обучения и достигают очень высокого удержания. оценки.
Таким образом, в соответствии с результатами по медоносным пчелам, опубликованными Pamir et al. (2011, 2014), из наших результатов мы пришли к выводу, что постепенно увеличивающаяся кривая обучения в среднем по группе не адекватно отражает поведение отдельных животных.Скорее, он смешивает три атрибута индивидуального обучения: способность или неспособность изучить данную задачу (учащиеся или не учащиеся), быстрое овладение правильным условным ответным поведением у учащихся и высокая устойчивость выражения условного ответа во время последовательные тренировки и попытки удержания памяти. Более того, мы могли бы получить такой же общий результат в задаче оперантного обучения таракана. Последний результат согласуется с исследованием на шмелях (Muller and Chittka, 2012), в котором наблюдалось, что одни люди всегда лучше других ассоциировали различные сигналы с наградой или наказанием в оперантной учебной задаче.
Интересно, что эти совпадающие результаты для медоносной пчелы и таракана, двух эволюционно далеко разделенных видов, противоречат давнему представлению о способностях к обучению у плодовых мух. Раннее сообщение об обонятельном обучении у Drosophila melanogaster , сделанное Quinn et al. (1974), используя хорошо зарекомендовавший себя и широко используемый групповой анализ классического обонятельного кондиционирования мух, пришел к выводу, что проявление поведения у особи было вероятностным, так что группу мух можно рассматривать как однородную в отношении способности приобретать правильное поведение CR. Это представление было поставлено под сомнение в более позднем исследовании (Chabaud et al., 2010), но оно требует дальнейшего убедительного исследования. Мы предполагаем, что дрозофилы демонстрируют индивидуальные способности к обучению, которые очень похожи на те, которые наблюдались у медоносных пчел и были установлены для тараканов в этом исследовании.
Это представление было поставлено под сомнение в более позднем исследовании (Chabaud et al., 2010), но оно требует дальнейшего убедительного исследования. Мы предполагаем, что дрозофилы демонстрируют индивидуальные способности к обучению, которые очень похожи на те, которые наблюдались у медоносных пчел и были установлены для тараканов в этом исследовании.
Возможные причины индивидуального выражения усвоенного поведения
Каковы могут быть основные причины наблюдаемой индивидуальности поведенческого обучения? На уровне нейронных цепей пластичность, вызванная обучением, наблюдалась в разных частях системы.Два исследования на медоносных пчелах обнаружили корреляцию между поведенческими характеристиками людей и проявлением пластичности нервной системы. Rath et al. (2011) выполнили визуализацию кальция в проекционных нейронах антеннальной доли. Для своего анализа они сформировали две подгруппы учащихся и не учащихся, основываясь на их условном ответном поведении, и сообщили, что в результате классического обонятельного кондиционирования паттерны реакции запаха в популяции проекционных нейронов стали более отчетливыми у учащихся, но не у других. учащиеся.Haenicke et al. (2018) выполнили Ca-визуализацию бутонов проекционных нейронов в чашечке грибовидного тела медоносной пчелы и обнаружили, что уровень нейрональной пластичности значительно коррелирует с уровнем поведенческой пластичности отдельных животных при классическом обонятельном кондиционировании. Было показано, что выходные нейроны грибовидного тела передают валентность запахов после классического кондиционирования пчел (Strube-Bloss et al., 2011, 2016) и мух (например, Aso et al., 2014; Hige et al., 2015).У пчел уровень наблюдаемой пластичности в этих нейронах после классического кондиционирования снова коррелирует с поведенческими характеристиками во время теста на удержание (Strube-Bloss, d’Albis, Menzel & Nawrot, неопубликованные данные). Таким образом, индивидуальность выполнения условных реакций во время сохранения памяти связана с лежащей в основе пластичностью нейронных схем.
Сообщалось о значительной корреляции между их чувствительностью к концентрации сахарозы и успеваемостью во время обонятельной задачи у пчел (Scheiner et al., 2004). Памир и др. (2014) повторно проанализировали данные Scheiner et al. (2001) показали, что чувствительность сахарозы, интерпретируемая как показатель состояния сытости, коррелирует с успеваемостью как в обонятельной, так и в тактильной классической обусловленности.
Кроме того, ряд исследований связали различия в способностях к обучению с генетическими вариациями у разных людей. Например, у медоносных пчел животные, которые хорошо справлялись с обонятельной / механосенсорной обработкой, также хорошо справлялись с визуальным обучением (Brandes and Menzel, 1990).С другой стороны, хорошие и плохие ученики из штаммов, отобранных для обонятельного кондиционирования, значительно различались по своим ценностям визуального обучения. Таким образом, между разными штаммами существуют генетические различия, и такая генетическая изменчивость может объяснять различия в обучении у людей (например, Brandes et al., 1988; Brandes and Menzel, 1990). Другое исследование медоносных пчел с учетом индивидуальных различий в задаче обучения латентному торможению (знание того, что некоторые стимулы являются , а не сигналами важных событий) также доказало генетическую предрасположенность к обучению этой задаче (Chandra et al., 2000). Кроме того, совсем недавнее исследование медоносных пчел показало, что генетический детерминизм лежит в основе компромисса между аппетитным и отталкивающим обучением (Junca et al., 2019). В другом исследовании плодовых мушек учили ассоциировать химический сигнал (хинин) с определенным субстратом. Это показало, что люди по-прежнему избегали этого субстрата через несколько часов после удаления сигнала, и ожидалось, что они внесут больше аллелей в следующее поколение. Примерно с 15 поколения экспериментальные популяции демонстрировали заметную способность избегать субстратов для откладки яиц, которые несколькими часами ранее содержали химический сигнал (см. Обзор: Mery and Kawecki, 2002; Dukas, 2008).В самом деле, генетическая изменчивость может лежать в основе индивидуальности поведения в целом и учебного поведения в частности. Однако, насколько нам известно, генетическая изменчивость тараканов не изучалась в зависимости от поведенческих черт. К сожалению, циклы созревания и размножения у тараканов довольно продолжительны.
Обзор: Mery and Kawecki, 2002; Dukas, 2008).В самом деле, генетическая изменчивость может лежать в основе индивидуальности поведения в целом и учебного поведения в частности. Однако, насколько нам известно, генетическая изменчивость тараканов не изучалась в зависимости от поведенческих черт. К сожалению, циклы созревания и размножения у тараканов довольно продолжительны.
Outlook
В настоящем исследовании мы исследовали индивидуальную успеваемость и скорость обучения в отдельных учебных задачах (классическое обонятельное кондиционирование или обучение оперантному месту).В будущих исследованиях мы расширим наш анализ индивидуальности в двух направлениях. Во-первых, мы исследуем, согласуется ли поведенческая характеристика людей в разных парадигмах обучения, то есть будут ли хорошие и быстрые ученики в одной классической парадигме обусловливания также работать выше среднего в различных классических или оперантных задачах кондиционирования. Насколько нам известно, существует только одно исследование беспозвоночных, в котором было опубликовано что-то сопоставимое с медоносными пчелами (Tait et al., 2019).Во-вторых, мы заинтересованы в согласованности в течение нескольких дней или недель, исследуя, является ли высокая / низкая производительность одного человека одинаково высокой / низкой при повторении той же или аналогичной задачи в более поздний момент времени.
Заявление о доступности данных
Наборы данных, созданные для этого исследования, доступны по запросу соответствующим авторам.
Авторские взносы
CA, JB и ND проводили эксперименты. CA, ND и MN разработали эксперименты и экспериментальные установки и написали рукопись.
Финансирование
CA получил стипендию от исследовательской группы обучения Анализ нейронных цепей на клеточном и субклеточном уровне , финансируемой Немецким исследовательским фондом (DFG-GRK 1960, грант № 233886668 MN). Дополнительное финансирование было получено от Немецкого исследовательского фонда в рамках исследовательского подразделения «Структура, пластичность и поведенческие функции грибовидного тела дрозофилы » (DFG-FOR 2705, грант № 403329959, выданный MN).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Сандру Мастани за вклад на ранней стадии этого проекта. Мы благодарим Питера Клоппенбурга за обсуждение, обмен средствами для борьбы с тараканами и использование животных. Мы благодарим команду нашего механического цеха под руководством Лео Лессона, который разработал и изготовил устройства для стимуляции запаха и гибкий набор для создания лабиринта для тараканов.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fphys.2019.01539/full#supplementary-material
Список литературы
Асо, Ю., Ситараман, Д., Ичиносе, Т., Каун, К. Р., Фогт, К., Беллиар-Герен, Г., и др. (2014). Выходные нейроны грибовидного тела кодируют валентность и направляют выбор действий на основе памяти у дрозофилы. elife 3, 1–42. DOI: 10.7554 / eLife.04580
CrossRef Полный текст | Google Scholar
Балдеррама, Н. (1980). Одно пробное обучение на американском таракане, Periplaneta americana . J. Insect Physiol. 26, 499–504. DOI: 10.1016 / 0022-1910 (80)
-7
CrossRef Полный текст | Google Scholar
Баррако Д. А., Ловелл К. Л. и Эйзенштейн Э. М. (1981). Влияние циклогексимида и пуромицина на обучение и удержание у таракана, P. americana . Pharmacol. Биохим. Behav. 15, 489–494. DOI: 10.1016 / 0091-3057 (81) -3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Белл, У. Дж., Бурк, Т., и Салдс, Г. Р. (1973). Феромон агрегации тараканов: направленная ориентация. Behav. Биол. 279, 251–255.
Google Scholar
Биттерман М. Э., Менцель Р., Фиц А. и Шефер С. (1983). Классическое кондиционирование удлинения хоботка у медоносных пчел ( Apis mellifera ). J. Comp. Psychol. 97, 107–119. DOI: 10.1037 / 0735-7036.97.2.107
J. Comp. Psychol. 97, 107–119. DOI: 10.1037 / 0735-7036.97.2.107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брандес, К., Фриш Б. и Мензель Р. (1988). Время формирования памяти различается у линий медоносных пчел, выбранных на хорошее и плохое обучение. Anim. Behav. 36, 981–985. DOI: 10.1016 / S0003-3472 (88) 80056-3
CrossRef Полный текст | Google Scholar
Брандес К. и Мензель Р. (1990). Общие механизмы кондиционирования удлинения хоботка и визуального обучения, выявленные генетическим отбором у медоносных пчел ( Apis mellifera capensis). J. Comp. Physiol. А 166, 545–552.DOI: 10.1007 / BF00192025
CrossRef Полный текст | Google Scholar
Камлитепе Ю., Аксой В. (2010). Первые свидетельства прекрасной способности различать цвета у муравьев (Hymenoptera, Formicidae). J. Exp. Биол. 213, 72–77. DOI: 10.1242 / jeb.037853
CrossRef Полный текст | Google Scholar
Carere, C., и Locurto, C. (2011). Взаимодействие между личностью животного и познанием животных. Curr. Zool. 57, 491–498. DOI: 10,1093 / czoolo / 57.4,491
CrossRef Полный текст | Google Scholar
Шабо, М.-А., Преат, Т., и Кайзер, Л. (2010). Поведенческая характеристика восстановления индивидуальной обонятельной памяти у Drosophila melanogaster . Фронт. Behav. Neurosci. 4, 1–11. DOI: 10.3389 / fnbeh.2010.00192
CrossRef Полный текст | Google Scholar
Чандра С. Б., Хослер Дж. С. и Смит Б. Х. (2000). Наследственная изменчивость латентного торможения и ее корреляция с обратным обучением у медоносных пчел ( Apis mellifera ). J. Comp. Psychol. 114, 86–97. DOI: 10.1037 / 0735-7036.114.1.86
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Читтка, Л., и Томсон, Дж. Д. (1997). Сенсорно-моторное обучение и его значение для специализации задач у шмелей. Behav. Ecol. Sociobiol. 41, 385–398. DOI: 10.1007 / s002650050400
DOI: 10.1007 / s002650050400
CrossRef Полный текст | Google Scholar
Дэвид М., Оклер Ю. и Сезилли Ф. (2011). Личность предсказывает социальное доминирование самок зебровых зябликов, Taeniopygia guttata , в контексте кормления. Anim. Behav. 81, 219–224. DOI: 10.1016 / j.anbehav.2010.10.008
CrossRef Полный текст | Google Scholar
Дингемансе, Н. Дж., И Вольф, М. (2010). Последние модели адаптивных личностных различий: обзор. Philos. Пер. R. Soc. Лондон. B Biol. Sci. 365, 3947–3958. DOI: 10.1098 / rstb.2010.0221
CrossRef Полный текст | Google Scholar
Дюпюи Ф., Сандос Ж.-К., Джурфа М. и Джозенс Р. (2006). Индивидуальное обонятельное обучение у муравьев Camponotus . Anim. Behav. 72, 1081–1091. DOI: 10.1016 / j.anbehav.2006.03.011
CrossRef Полный текст | Google Scholar
Эрбер, Дж., Приббенов, Б., Гранди, К., и Кежек, С. (1997). Тактильное моторное обучение в системе усиков медоносной пчелы ( Apis mellifera L.). J. Comp. Physiol. А 181, 355–365. DOI: 10.1007 / s0035
121
CrossRef Полный текст | Google Scholar
Галлистель, К. Р., Фэрхерст, С., и Бальзам, П. (2004). Кривая обучения: последствия количественного анализа. Proc. Natl. Акад. Sci. USA 101, 13124–13131. DOI: 10.1073 / pnas.0404965101
CrossRef Полный текст | Google Scholar
Джурфа М., Хаммер М., Стах С., Столлхофф Н., Мюллер-Дейсиг Н. и Мизирицки К. (1999). Паттерн обучения пчелами: процедура кондиционирования и стратегия распознавания. Anim. Behav. 57, 315–324. DOI: 10.1006 / anbe.1998.0957
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джурфа М. и Сандос Дж.-C. (2012). Обучение и память беспозвоночных: пятьдесят лет обонятельного кондиционирования реакции расширения хоботка у медоносных пчел. ЖЖ. Mem. 19, 54–66. DOI: 10.1101 / lm.024711.111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джурфа М. , Чжан С., Дженетт А., Мензель Р. и Сринивасан М. В. (2001). Понятия «одинаковость» и «различие» у насекомых. Природа 410, 930–933. DOI: 10.1038 / 35073582
, Чжан С., Дженетт А., Мензель Р. и Сринивасан М. В. (2001). Понятия «одинаковость» и «различие» у насекомых. Природа 410, 930–933. DOI: 10.1038 / 35073582
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гослинг, С.Д., и Вазир, С. (2002). Мы лаем на правильное дерево? Сравнительный подход к оценке личности. J. Res. Чел. 36, 607–614. DOI: 10.1016 / S0092-6566 (02) 00511-1
CrossRef Полный текст | Google Scholar
Гринстед, Л., Прюитт, Дж. Н., Сеттепани, В., и Билде, Т. (2013). Индивидуальные личности формируют дифференциацию задач у социального паука. Proc. R. Soc. B Biol. Sci. 280: 20131407. DOI: 10.1098 / rspb.2013.1407
CrossRef Полный текст | Google Scholar
Хенике, Дж., Ямагата, Н., Звака, Х., Наврот, М., и Мензель, Р. (2018). Нейронные корреляты обучения запахам в пресинаптических микрогломерулярных цепях чашечки грибовидного тела медоносной пчелы. eNeuro 5: ENEURO.0128-18.2018. DOI: 10.1523 / ENEURO.0128-18.2018
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хиге Т., Асо Ю., Рубин Г. М. и Тернер Г. К. (2015). Индивидуализация обонятельного кодирования в выходных нейронах грибовидного тела, обусловленная пластичностью. Природа 526, 258–262.DOI: 10.1038 / nature15396
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ходжес, К. М., Стейдл, Дж. Л. М., Веррен, Дж. Х., Вет, Л. Е. М. и Смид, Х. М. (2012). Высокопроизводительный тест на обонятельное кондиционирование и тест на сохранение памяти показывают вариабельность у паразитических ос Nasonia. Genes Brain Behav. 11, 879–887. DOI: 10.1111 / j.1601-183X.2012.00823.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хосоно, С., Мацумото, Ю., Мизунами, М.(2016). Взаимодействие тормозящего и облегчающего эффектов условных проб на формирование долговременной памяти. ЖЖ. Mem. 23, 669–678. DOI: 10.1101 / лм. 043513.116
043513.116
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Junca, P., Garnery, L., and Sandoz, J.-C. (2019). Генотипический компромисс между аппетитными и отталкивающими способностями у медоносных пчел. Sci. Реп. 9, 1–14. DOI: 10.1038 / s41598-019-46482-4
CrossRef Полный текст | Google Scholar
Колата, С., Лайт, К., Таунсенд, Д. А., Хейл, Г., Гроссман, Х. К., и Мацель, Л. Д. (2005). Вариации в объеме рабочей памяти предсказывают индивидуальные различия в общих способностях к обучению генетически разнородных мышей. Neurobiol. Учиться. Mem. 84, 241–246. DOI: 10.1016 / j.nlm.2005.07.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Kralj-Fišer, S., and Schuett, W. (2014). Изучение изменчивости личности у беспозвоночных: зачем беспокоиться? Anim. Behav. 91, 41–52. DOI: 10.1016 / j.anbehav.2014.02.016
CrossRef Полный текст | Google Scholar
Кувабара, М. (1957). Bildung des bedingten Reflexes von Pavlovs Typus bei der Honigbiene, Apis mellifica. J. Fat. Sci. Hokkaido Univ. 13, 458–464.
Google Scholar
Квон, Х.-В., Пост, Д. Д., и Штраусфельд, Н. Дж. (2004). Пространственное обучение у сдержанного американского таракана Periplaneta americana . J. Exp. Биол. 207, 377–383.DOI: 10.1242 / jeb.00737
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лаупрасерт П., Лаупрасет П., Ситтичароенчай Д., Тиракхупт К. и Прадацударасар А.-О. (2006). Пищевые предпочтения и пищевое поведение немецкого таракана, Blattella germanica (Linnaeus). J. Sci. Res. Chula Univ. 31, 121–126. Доступно по адресу: http://www.thaiscience.info/journals/Article/CJSR/10324268.pdf
Google Scholar
Пост, Д.Д., Квон, Х.-В. (2004). Движение антенн свидетельствует об ассоциативном обучении американского таракана Periplaneta americana . J. Exp. Биол. 207, 369–375. DOI: 10.1242 / jeb.00736
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю, Ж.-Л., и Сакума, М. (2013). Обонятельное кондиционирование с помощью отдельных химических веществ у немецкого таракана, Blattella germanica (Dictyoptera: Blattellidae). Заявл. Энтомол. Zool. 48, 387–396.DOI: 10.1007 / s13355-013-0199-x
CrossRef Полный текст | Google Scholar
Ланни, Г. Х. (1970). Использование дисперсионного анализа с дихотомической зависимой переменной: эмпирическое исследование. J. Educ. Измер. 7, 263–269. DOI: 10.1111 / j.1745-3984.1970.tb00727.x
CrossRef Полный текст | Google Scholar
Menzel, R., Durst, C., Erber, J., Eichmüller, S., Hammer, M., Hildebrandt, H., et al. (1994). Грибовидные тела пчелы: от молекул к поведению. Fortschr. Zool. 39, 81–102.
Google Scholar
Mery, F., and Kawecki, T. J. (2002). Экспериментальная эволюция обучаемости у плодовых мушек. Proc. Natl. Акад. Sci. USA 99, 14274–14279. DOI: 10.1073 / pnas.222371199
CrossRef Полный текст | Google Scholar
Мизунами М., Вейбрехт Дж. М. и Штраусфельд Н. Дж. (1998). Грибные тела таракана: их участие в памяти места. J. Comp. Neurol. 402, 520–537.DOI: 10.1002 / (SICI) 1096-9861 (19981228) 402: 4 <520 :: AID-CNE6> 3.0.CO; 2-K
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мюллер, Х., Читтка, Л. (2012). Стабильные межиндивидуальные различия в способностях шмелей различать по цвету, форме и запаху (перепончатокрылые: Bombus terrestris ). Энтомол. Gener. 34, 1–6. DOI: 10.1127 / entom.gen / 34/2012/1
CrossRef Полный текст | Google Scholar
Нувиан, М., и Галиция, К. Г. (2019). Аверсивная тренировка медоносных пчел в автоматизированном Y-лабиринте. Фронт. Physiol. 10, 1–17. DOI: 10.3389 / fphys.2019.00678
CrossRef Полный текст | Google Scholar
Памир Э., Чакроборти Н. К., Столлхофф Н., Геринг К. Б., Антеманн В., Моргенштерн Л. и др. (2011). Среднее групповое поведение не отражает индивидуального поведения в классическом кондиционировании медоносной пчелы. ЖЖ. Mem. 18, 733–741. DOI: 10.1101 / лм. 2232711
К., Столлхофф Н., Геринг К. Б., Антеманн В., Моргенштерн Л. и др. (2011). Среднее групповое поведение не отражает индивидуального поведения в классическом кондиционировании медоносной пчелы. ЖЖ. Mem. 18, 733–741. DOI: 10.1101 / лм. 2232711
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Памир, Э., Шишка П., Шайнер Р. и Наврот М. П. (2014). Динамика быстрого обучения у отдельных медоносных пчел во время классического кондиционирования. Фронт. Behav. Neurosci. 8: 313. DOI: 10.3389 / fnbeh.2014.00313
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Павлов И. П. (1927). Условные рефлексы. Исследование физиологической активности коры головного мозга . Оксфорд, Англия: Издательство Оксфордского университета.
Google Scholar
Пинтер-Уоллман, Н.(2012). Личность у социальных насекомых: как личность рабочего определяет личность колонии? Curr. Zool. 58, 580–588. DOI: 10.1093 / czoolo / 58.4.580
CrossRef Полный текст | Google Scholar
Planas-Sitjà, I., and Deneubourg, J.-L. (2018). Роль изменчивости личности, пластичности и социальной поддержки в агрегации тараканов. Biol. Откройте 7: bio036582. DOI: 10.1242 / bio.036582
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Planas-Sitjà, I., Nicolis, S.C., Sempo, G., and Deneubourg, J.-L. (2018). Взаимодействие между личностями и социальными взаимодействиями влияет на сплоченность группы и скорость агрегации. PLoS One 13: e0201053. DOI: 10.1371 / journal.pone.0201053
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Pomaville, M. B., and Lent, D. D. (2018). Множественные изображения космоса тараканом, Periplaneta americana . Фронт. Psychol. 9, 1–15.DOI: 10.3389 / fpsyg.2018.01312
CrossRef Полный текст | Google Scholar
Куинн, В. Г., Харрис, В. А., и Бензер, С. (1974). Обусловленное поведение у Drosophila melanogaster (обучение / память / распознавание запахов / цветовое зрение). Proc. Natl. Акад. Sci. USA 71, 708–712.
Google Scholar
Рат, Л., Джованни Галиция, К., и Шишка, П. (2011). Множественные следы памяти после ассоциативного обучения в антеннальной доли медоносной пчелы. Eur.J. Neurosci. 34, 352–360. DOI: 10.1111 / j.1460-9568.2011.07753.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роуз Дж., Каллен Д. А., Симпсон С. Дж. И Стивенсон П. А. (2017). Рожденные побеждать или воспитанные проигрывать: агрессивные и покорные поведенческие профили сверчков. Anim. Behav. 123, 441–450. DOI: 10.1016 / j.anbehav.2016.11.021
CrossRef Полный текст | Google Scholar
Сакура, М., Мизунами, М. (2001). Обонятельное обучение и память у таракана Periplaneta americana . Zool. Sci. 18, 21–28. DOI: 10.2108 / zsj.18.21
CrossRef Полный текст | Google Scholar
Сакура М., Окада Р. и Мизунами М. (2002). Обонятельная дискриминация тараканами близких по структуре спиртов. J. Comp. Physiol. Нейроэтол. Sens. Neural Behav. Physiol. 188, 787–797. DOI: 10.1007 / s00359-002-0366-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шайнер Р., Пейдж Р. Э. и Эрбер Дж. (2001). Влияние генотипа, кормовой роли и отзывчивости сахарозы на тактильное обучение медоносных пчел ( Apis mellifera L.). Neurobiol. Учиться. Mem. 76, 138–150. DOI: 10.1006 / NLME.2000.3996
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шайнер Р., Пейдж Р. Э. и Эрбер Дж. (2004). Отзывчивость сахарозы и пластичность поведения у медоносных пчел ( Apis mellifera ). Apidologie 35, 133–142. DOI: 10.1051 / apido: 2004001
CrossRef Полный текст | Google Scholar
Шуберт М., Ханссон Б. С. и Саксе С. (2014). Банановый код – естественная обработка смеси в обонятельной схеме Drosophila melanogaster . Фронт. Physiol. 5, 1–13. DOI: 10.3389 / fphys.2014.00059
CrossRef Полный текст | Google Scholar
Шуэтт В. и Далл С. Р. X. (2009). Половые различия, социальный контекст и личность у зебровых зябликов, Taeniopygia guttata . Anim. Behav. 77, 1041–1050. DOI: 10.1016 / j.anbehav.2008.12.024
CrossRef Полный текст | Google Scholar
Schuett, W., Dall, S.R.X., Baeumer, J., Kloesener, M.H., Nakagawa, S., Beinlich, F., и другие. (2011). Изменчивость личности клонального насекомого: гороховая тля, Acyrthosiphon pisum . Dev. Psychobiol. 53, 631–640. DOI: 10.1002 / dev.20538
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сих А., Белл А. и Джонсон Дж. К. (2004a). Поведенческие синдромы: экологический и эволюционный обзор. Trends Ecol. Evol. 19, 372–378. DOI: 10.1016 / j.tree.2004.04.009
CrossRef Полный текст | Google Scholar
Сих, А., Белл, А. М., Джонсон, Дж. К., и Зиемба, Р. Э. (2004b). Поведенческие синдромы: интегративный обзор. Q. Rev. Biol. 79, 241–277. DOI: 10.1086 / 422893
CrossRef Полный текст | Google Scholar
Strube-Bloss, M. F., Nawrot, M. P., and Menzel, R. (2011). Выходные нейроны грибовидного тела кодируют ассоциации “запах-вознаграждение”. J. Neurosci. 31, 3129–3140. DOI: 10.1523 / JNEUROSCI.2583-10.2011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Штрубе-Блосс, М.Ф., Наврот М. П., Мензель Р. (2016). Нейронные корреляты побочной памяти запахов в выходных нейронах грибовидного тела. Proc. R. Soc. B Biol. Sci. 283: 2016 1270. DOI: 10.1098 / rspb.2016.1270
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тейт К., Маттисе-Лоренцен А., Ларк А. и Науг Д. (2019). Индивидуальные различия в обучаемости пчел. Behav. Процесс. 167: 103918. DOI: 10.1016 / j.beproc.2019.103918
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такеда, К.(1961). Классическая условная реакция у медоносной пчелы. J. Insect Physiol. 6, 168–179. DOI: 10.1016 / 0022-1910 (61)
-9
CrossRef Полный текст | Google Scholar
Тернер, К. Х. (1912). Экспериментальное исследование очевидной обратной реакции на свет у плотвы (Periplaneta Orientalis L.). 23, 371–386.
Google Scholar
Ватанабэ, Х., Кобаяси, Ю., Сакура, М., Мацумото, Ю., и Мизунами, М. (2003). Классическое обонятельное кондиционирование у тараканов Periplaneta americana . Zool. Sci. 20, 1447–1454. DOI: 10.2108 / zsj.20.1447
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райт, К. М., Холбрук, К. Т., и Прюитт, Дж. Н. (2014). Личность животного совмещает специализацию задачи и мастерство выполнения задачи в обществе пауков. Proc. Natl. Акад. Sci. USA 111, 9533–9537. DOI: 10.1073 / pnas.1400850111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Динамика эмиграции тараканов при различных режимах возмущения не зависит от индивидуальных особенностей
Влияние личности и размера группы во время активной фазы и фазы покоя
Группы из 20 тараканов тестировались на арене с двумя одинаковыми укрытиями в течение недели (см. Методы) .В контрольном или ненарушенном состоянии (UD) убежища оставались нетронутыми, тогда как в состоянии с частыми нарушениями (D) выбранный укрытие для тараканов в первый день ежедневно тревожили светом. Во-первых, мы проанализировали последовательность поведения тараканов в активной фазе (ночное время), когда они исследовали арену. С этой целью мы приняли во внимание различные параметры, которые были описаны ранее как хорошие показатели использования укрытий и кормодобывающей деятельности. Тараканы показали значительную стабильность поведения по всем исследованным параметрам (таблица 1) в условиях UD и D.Действительно, коэффициенты Кендалла, полученные для Общего времени вне (TTO) и других временных показателей, таких как продолжительность первого выхода, были больше, чем ожидалось случайным распределением Кендалла (KRD), которое предполагает, что все люди идентичны. Высокие значения коэффициента Кендалла в диапазоне от 0 до 1 указывают на согласованность между днями (см. Методы).
Таблица 1 Сводка значений W. Кендалла.
Кроме того, мы проанализировали, имела ли место различная социальная динамика внутри групп.Для этого мы создали искусственные или смоделированные группы из экспериментальных данных. Каждая смоделированная группа состояла из 20 человек, случайно выбранных из пула данных тестируемых лиц (120 для состояния D и 140 для состояния UD). Используя этот метод, люди из искусственных групп сохраняли последовательность своего поведения (собственный TTO в течение 4 ночей), но были смешанными между группами. Мы сравнили сумму TTO индивидуумов в экспериментальных группах с суммой TTO искусственных групп (рис.1), а между распределениями не наблюдалось значительных различий (тест KS: D 52,200 = 0,26462, P = 0,3014), что указывало на отсутствие разной социальной динамики внутри групп, поскольку смешивание TTO отдельных лиц не влияло на группы. Распределение TTO. Если бы группы демонстрировали разную социальную динамику, мы бы наблюдали изменения в распределении ВПД в группах после смешивания людей из разных групп.
Рис. 1
Сравнение между TTO групп экспериментальных групп (серые столбцы) и TTO групп искусственных групп (красная линия).
Что касается фазы покоя (дневное время), мы проанализировали влияние размера группы во время беспорядков, и корреляции между временем, необходимым для выхода из убежища S после нарушения, и размером группы в начале нарушения не наблюдалось. (регрессионный тест для первого нарушения каждого дня: R 2 = 0,01, F 1,233 = 73,63, P = 0,58), который показал, что влияние размера группы на эмиграционное поведение незначительно. Процесс привыкания не наблюдался, и после большего количества возмущений у особей не уменьшалось время реакции.Действительно, среднее индивидуальное время, необходимое для выхода из укрытия во время нарушений, не коррелировало с количеством нарушений, воздействующих на одного и того же человека (корреляционный тест Кендалла: R = 0,06, τ = 0,89, N = 113, P = 0,37).
Моделирование динамики эмиграции и стабильности выбора в нарушенных и ненарушенных условиях
Мы предлагаем стохастическую модель с целью прогнозирования поведения тараканов во времени и проверки того, воспроизводит ли группа идентичных людей без индивидуальной памяти глобальную динамику эмиграции и поведенческие модели. дисперсия ответа.Следовательно, модели нужна средняя вероятность выхода из убежища и присоединения к нему во время активной фазы и фазы отдыха. С этой целью мы использовали экспериментальные результаты для количественной оценки доли особей из всех групп (состояние UD и D отдельно), которые находились в убежище в начале данной фазы и переехали в другое убежище в конце ее (активное или фаза покоя). Мы рассчитали эти средние вероятности для выбранных (S) и невыбранных (NS) укрытий на обоих этапах и реализовали их в нашей модели.Исходное состояние для этой модели было асимметричным, что было таким же, как и в наших наблюдениях, потому что большинство тараканов находились под укрытием S. в начале экспериментов. Та же модель была выполнена для условий UD и D с соответствующими значениями вероятности (см. Рис. 2). Кроме того, для условия D мы показываем, что TTO человека не влияет на переходы между убежищами. Другими словами, особи, переходящие из одного убежища в другое или в одно и то же во время активной фазы, не показали существенно различающихся TTO (см. Таблицу 2).Что касается условия UD, из-за большого количества случаев между S и S укрытие от других возможных переходов (S в NS, NS в NS и NS в S), мы не смогли выполнить какой-либо надежный статистический тест.
Рис. 2
Схема, представляющая вероятности посещения определенного убежища во время активной и неактивной фаз: ( a ) Ненарушенное состояние; и ( b ) Нарушенное состояние.
Таблица 2 Значения P для сравнения экспериментальных распределений TTO среди всех переходов между убежищами S и NS в состоянии D (критерий Манна-Уитни).
На рис. 2 показана диаграмма, основанная на наших результатах, которая описывает пространственную динамику тараканов в течение 24 часов (покой + активная фаза) для состояний UD и D. Поскольку TTO не влияет на переход между убежищами, эта модель не учитывает ночные действия (например, TTO) и основана только на индивидуальных вероятностях перехода между убежищами. Что касается состояния UD (рис. 2а), тараканы никогда самопроизвольно не покидали убежища во время фазы покоя; поэтому единственное перемещение между укрытиями происходило во время активной фазы.Во время этой фазы существовала вероятность перехода к противоположному укрытию, причем Ψ (S, NS) и Ψ (NS, S) представляли вероятности перехода от S к убежищу NS и от NS к укрытию S. ночь соответственно. Кроме того, Ψ (NS, NS) (= 1 − Ψ (NS, S)) и Ψ (S, S) (= 1 − Ψ (S, NS)) – это вероятности возврата в ранее занятое убежище. В нашем экспериментальном случае индивидуальная вероятность Ψ (NS, S) была больше, чем ее противоположность, что означает, что убежище S было более привлекательным, чем укрытие NS (Ψ (S, NS) = 0.1; Ψ (NS, S) = 0,4, тест KS: D 1,3 = 1, P = 0,01; Рис. 2а).
Что касается условия D (рис. 2b), схема становится более сложной из-за того, что тараканы перемещаются между укрытиями во время фазы покоя. Во время активной фазы тараканы могли перемещаться между убежищами S и NS, что было аналогично состоянию UD, хотя эти вероятности для состояния D существенно не различались (Ψ (NS, S) = 0,3 и Ψ (S , NS) = 0,2, тест KS: D 1,3 = 0.5, P = 0,53). Сравнение обоих условий показало, что Ψ (NS, S) существенно не различались (тест KS: D 1,3 = 0,5, P = 0,53). Тем не менее, (S, NS) для условия D было больше, чем для условия UD (тест KS: D 1,3 = 1, P = 0,01, см. Рис. 2). Что касается фазы отдыха, то особи в приюте NS не двигались самопроизвольно, как в состоянии UD. Тем не менее, тараканы в убежище S имели вероятность перебраться в укрытие NS, если их потревожили.Вероятность переехать (Δ (S, NS)) и остаться в убежище S (Δ ( S, S )) к концу фазы покоя определяется уравнениями (1) и (2) соответственно. В состоянии UD Δ ( S, NS ) = 0; а в условии D Δ ( S, NS ) = 0,1. В этом случае (уравнения 1-2) θ – это условная вероятность перемещения между южным и южным укрытиями для каждого нарушения, а l – количество нарушений в день.
Экспериментальная эволюция количества тараканов, поселившихся внутри каждого убежища в условиях UD и D, воспроизводится с помощью моделирования и уравнений (3–5) (сравнение с наблюдаемыми результатами: тест KS: D 1,4 = 0.4, P = 0,7). Уравнения (3–5) рассматривают различные вероятности перехода между убежищами S и NS. P (n ) ( Q (n )) – вероятность заселения тараканов на фазу отдыха в убежище S (NS) в день n и зависит от вероятностей передвижения во время фазы отдыха и активной фазы. Tr – это результирующая вероятность остаться в убежище S, как n -1, и связана с реакциями беспокойства и ночной активностью тараканов.
Решение уравнений (3–5) выглядит следующим образом:
Стационарное состояние задается уравнением. (7):
В состоянии D из-за суточных нарушений численность населения в убежище S уменьшается каждый день, а популяция в убежище NS увеличивается, пока в конце недели оно не достигнет значения, близкого к стационарному состоянию. (= 0,54), предсказанное моделью через 5 дней (см. Уравнение 7 и рис. 3a). Кроме того, ур. (7) предсказывает, что большие значения Δ (S, NS) снижают вероятность проживания в убежище S в стационарном состоянии, что означает, что окончательная доля укрытых тараканов в убежище S будет близка к Ψ (NS, S ).В состоянии UD, в котором отсутствует суточная миграция (Δ (S, NS) = 0), Tr равно Ψ (S, S). Интересно, что средняя доля тараканов, поселившихся в убежище S в начале эксперимента, уже была близка к предсказанному стационарному состоянию, предсказанному моделью в состоянии UD (= 0,8). Это соглашение предполагает, что без возмущений стационарное состояние быстро достигается с первого дня.
Рис. 3: Сравнение наших наблюдений и теоретических результатов.
( a ) Наблюдаемая и теоретическая средняя доля лиц, укрытых внутри каждого убежища на протяжении всего периода исследования в течение дня (± стандартное отклонение). Частота количества дней, в течение которых каждый человек находился под убежищем Selected (S) в ( b ) Беспокойном и ( c ) Непотревоженном состоянии. ( d ) Частота общего количества нарушений, испытываемых человеком.
Кроме того, мы использовали уравнения (8–9) для оценки количества особей в убежище S в начале каждой фазы отдыха.Вероятность заселения в убежище S или NS ( P (n, i ) и Q (n, i ) соответственно) в начале дня n и с i предыдущих дней в S Укрытие дается уравнениями (8–9).
Теоретическое распределение полной вероятности ( P (n, i ) + Q (n, i )) аналогично экспериментальным результатам для условий UD и D (тест KS: D 1,5 = 0,17, P = 1 и тест KS: D 1,5 = 0.33, P = 0,93 соответственно, рис. 3б, в). Для обоих условий количество тараканов, которые проводят от 5 до 0 дней под выбранным убежищем, непрерывно уменьшается (рис. 3b, c). Отсутствие различий между нашей моделью, которая рассматривает идентичных людей, и нашими экспериментальными частотами убедительно свидетельствует о том, что личность не повлияла на верность сайта.
Для условия D мы проанализировали общее количество нарушений, испытываемых каждым человеком (рис. 3d). Действительно, большинство тараканов, отдыхающих в убежище S, испытывали все 6 нарушений в день.Вероятность заселения в убежище S ( P E ( n, k )) или укрытие NS ( Q E ( n, k )) в конце дня n после возникновения k возмущений выглядит следующим образом (см. SI 1 для подробного объяснения):
Теоретические результаты, полученные с помощью уравнений (10–12), и наблюдаемые возмущения, испытываемые каждым человеком (рис. 3d), коррелированы . Более того, теоретическое распределение количества возмущений, возникающих в конце каждой фазы покоя, не отличается от экспериментального распределения (тест KS: D 1,5 = 0.5, Р = 0,44; см. SI 2).
Тараканы могут учиться – как собаки и люди
ГОНКОНГ (Рейтер) – Тараканы обладают памятью, и их можно научить выделять слюну в ответ на нейтральные раздражители так же, как собаки Павлова, когда знаменитый русский врач позвонил в свой звонок. , Выяснили японские исследователи.
Гигантских мадагаскарских шипящих тараканов видели в Бангкоке 23 мая 2003 года. У тараканов есть память, и их можно научить выделять слюну в ответ на нейтральные раздражители так же, как собаки Павлова, когда знаменитый русский доктор позвонил в свой звонок, японские исследователи нашел.REUTERS / Sukree Sukplang
Такая «обусловленность» может иметь место только при наличии памяти и обучения, а такая реакция выделения слюны ранее была доказана только на людях и собаках.
Кажется, тараканы тоже обладают такой способностью.
В последнем выпуске онлайн-журнала Public Library of Science исследователи заявили, что надеются узнать больше о человеческом мозге, исследуя то, что происходит в более простом мозге таракана. (Статья находится в свободном доступе здесь)
«Понимание механизма обучения мозга насекомых может помочь нам понять функции человеческого мозга.«Есть много, много общих характеристик», – сказал Макото Мизунами из Высшей школы естественных наук Университета Тохоку в телефонном интервью.
В ходе эксперимента ученые подвергали группу тараканов запаху всякий раз, когда они кормили их раствором сахара. Они обнаружили, что когда они позже подвергали тараканов воздействию одного запаха, они все еще пускали слюни.
Другой группе тараканов кормили сахарным раствором без запаха, и последующее воздействие запаха не привело к изменению количества выделяемой слюны.
«Конечно, тараканы могут запоминать и учиться», – сказал Мизунами.
В 1890-х и 1990-х годах русский врач Иван Петрович Павлов проводил исследования того, что сейчас известно как «классическая обусловленность» с собаками.
Он использовал колокольчики, чтобы позвать собак к еде, и после нескольких повторений у собак началось выделение слюны в ответ на звонок.
Но до сегодняшнего дня, несмотря на достижения науки, мало что известно о мозге млекопитающих и его нервных механизмах, потому что они очень сложны.
Мизунами сказал, что его команда выяснит, что происходит в мозгу таракана в следующий раз.
«В мозгу таракана очень много нейронов, и мы должны найти, какой нейрон на самом деле отвечает за это обучение», – сказал он.
«Мы считаем, что очень важно изучить простую систему, чтобы очень точно определять, что происходит во время обучения. Мы надеемся, что это, по крайней мере, частично применимо к людям ».
Абсурдное существо недели: Оса, которая порабощает тараканов укусом в мозг
Оказавшись в норе, оса бесцеремонно запихивает таракана внутрь и кладет одно яйцо на его ногу, после чего начинает собирать камешки.Этим она закапывает свою жертву и свое прожорливое потомство. Вот, посмотрите немного на моего ребенка , кажется, она говорит.
«Это занимает около 30 минут, в течение которых таракан не пытается вырвать яйцо из своей ноги или выбраться из своей (пока еще широко открытой) гробницы», – сказал Гал. «Когда оса завершает запечатывание норы, что призвано защитить тараканов (а также яйцо и развивающуюся личинку) от потенциальных хищников, а не убежать, она улетает в поисках новой добычи.
Примерно через два дня из яйца вылупляется личинка, прогрызая отверстие в брюшке таракана, питаясь гемолимфой, которая сочится наружу. Затем он забирается в брюшную полость, как Люк пинает его в таунтауне, хотя, в отличие от Люка, личинка поедает органы, ловко сохраняя нервную систему напоследок. Как только это исчезнет, таракан наконец умирает, и в этот момент личинка выделяет антимикробное соединение на стенках теперь уже пустого насекомого. Он окуклится в этом продезинфицированном (в конце концов, это труп) доме в течение месяца, а затем, наконец, прорвется сквозь шелуху, как взрослая оса.
Как и люди, детенышей жемчужной осы рожают аисты, в данном случае выдолбленный труп таракана.
Изображение: Ram Gal
Roach Coached
Так что же тогда было в том яде, который мог так тщательно зомбировать таракана? Гэл еще не уверена, за исключением того, что это смесь соединений, и похоже, что один и тот же яд используется для укуса ног и мозга. «Поэтому мы предполагаем, что различные компоненты в этом коктейле ядов и их различное воздействие на разные части нервной системы таракана ответственны за различные поведенческие манипуляции (кратковременный паралич, уход за телом и« зомбификацию »).
Интересно, что если Гал удалит яйцо из ноги зомбированного таракана в лаборатории, оно вернется к нормальному поведению через несколько дней. По мнению Гал, это говорит о том, что зомбификация – более тонкая манипуляция, чем полный захват нервной системы таракана. Кроме того, помните, что таракан будет ходить, как обычно, и поправится, как обычно, если вы его перевернете, демонстрируя, что сенсорные и моторные функции в норме.
Тем не менее, он хочет добровольно следовать оса его гибели, так что мы имеем здесь на самом деле загадка свободной воли – это весьма спорный вопрос вы достаточно соответствующим образом решили не обратить внимание на в философии 101 лекций (Rush сделал довольно sicknasty Версия CliffsNotes, если вам нравится вся краткость).Таракан вполне способен убежать, так почему же нет? «Это означает, что, как и млекопитающие, насекомые – это не просто« автоматы », которые детерминированно реагируют на внешние раздражители (распространенное заблуждение как среди ученых, так и среди непрофессионалов), – сказал Гал, – но вместо этого имеют базовую способность выбирать, когда действовать. и какие действия предпринять ».
Охранные судебные приказы и запретительные судебные приказы – RAM Law PLLC
Защитные судебные приказы
Защитный судебный приказ, обычно называемый «OFP», представляет собой запретительный судебный приказ, на который можно подать прошение, чтобы остановить насилие в семье.Человек может подать в суд ходатайство о выдаче постановления о защите в случае совершения домашнего насилия. Лицо, обращающееся в суд с просьбой о предоставлении защиты, называется истцом, а предполагаемый обидчик – ответчиком. Существуют различные действия, которые могут представлять собой домашнее насилие с целью получения судебного приказа о защите, включая нанесение физического вреда, угрозу физического вреда, принуждение к сексуальным контактам или вмешательство в вызов службы экстренной помощи. Сексуальное насилие в отношении несовершеннолетнего также может быть основанием для вынесения судом постановления о защите.
Приказ о защите, в случае его выдачи, предотвращает контакты между предполагаемым обидчиком и членами семьи или домохозяйства. Приказ о защите специально разработан для предотвращения продолжающегося домашнего насилия, потому что он применяется только к делам, связанным с супругом или бывшим супругом, родителями и детьми, теми, кто живет вместе или ранее проживал вместе, теми, кто связан кровными связями или имеет одного ребенка, тем, кто вовлечен в романтические или сексуальные отношения. Поскольку он предназначен для предотвращения домашнего насилия, судебный приказ о защите может означать, что предполагаемый обидчик не может вернуться в свой дом, связаться со своими детьми или вообще общаться со своим супругом или другим значимым лицом.Нарушение судебного приказа о защите является преступлением, и любой, кто получил приказ о защите от него, должен немедленно проконсультироваться с адвокатом по уголовным делам для выяснения своих прав.
Запретительный ордер на преследование
Хотя судебный приказ о защите может быть выдан только при наличии семейных или домашних отношений между заявителем и предполагаемым обидчиком, запретительный ордер на преследование (обычно известный как СПЧ) не содержит этого требования.